Взаимодействие в лимфоцита и т лимфоцита хелпера

Варианты взаимодействия $T$- и $B$-лимфоцитов при иммунном ответе
Единственными эффективными антигенпрезентирующими клетками при первичном иммунном ответе для $T$-лимфоцитов являются дендритные клетки. Если при активации $T$-лимфоцита антиген был презентирован дендритными клетками, то в иммунный ответ вовлекаются $B$-лимфоциты, расположенные рядом.
Возможны следующие варианты взаимодействия $T$- и $B$-лимфоцитов:
- $B$-лимфоциты связывают растворимый антиген через BCR, путем эндоцитоза его поглощают, подвергают процессингу и на поверхности экспонируют фрагменты антигена совместно с молекулами МНС-II. Антиген связывается на поверхности $B$-лимфоцита TCR в комплексе с МНС-II. Устанавливаются необходимые корецепторные связи между $T$- и $B$-лимфоцитами. Такое взаимодействие наблюдается в $T$-зависимых зонах периферической лимфоидной ткани в самом начале развития иммунного ответа.
- Специфический антиген распознается $B$-лимфоцитом и вблизи находится активированный $T$-лимфоцит, который распознал антиген на другой антигенпрезентирующей клетке. Взаимодействие $T$- и $B$-лимфоцитов может проявиться восприятием цитокиновых сигналов $B$-клеткой от $B$-лимфоцита.
Замечание 1
Взаимодействие с участием мембранных молекул при первичном иммунном ответе может наступать или нет. При вторичном иммунном ответе принимают участие $T$-клетки памяти. В обязательном порядке происходит взаимодействие мембранной молекулы $B$-лимфоцита $CD40 с$ мембранной молекулой $T$-клетки $CD40L$. Вторичный ответ $B2$-лимфоцитов происходит с обязательным переключением изотипа иммуноглобулинов с $IgM$ на $Iga$, $IgG$, $IgE$. Эти взаимодействия происходят в фолликулах лимфоидных органов (в $B$-клеточных зонах).
Реализация реакции иммунной системы на антиген
После внедрения антигена во внутреннюю среду организма наблюдается его фагоцитирование или поглощение макрофагами. Макрофаги с помощью протеолитических ферментов убирают детерминантные группировки чужих макромолекул. Образовавшиеся детерминанты в результате экзоцитоза попадают на поверхность мембраны, где соединяются с мембранными белками макрофага.
Белки макрофага кодируются генами главного комплекса гистосовместимости, которые обеспечивают синтез белков, определяющих иммунологическую индивидуальность организма.
Антигены гистосовместимости (мембранные белки) являются трансплантационными антигенами, так как различны у разных индивидов. Эти антигены вызывают реакцию отторжения трансплантата при пересадке тканей и органов.
Антигены гистосовместимости представляют $T$-лимфоцитам очищенную антигенную детерминанту.
Передача антигена с макрофага $T$-лимфоциту происходит в результате следующих процессов:
- $T$-лимфоцит определяет чежеродность детерминантной группы антигена, находящегося на поверхности макрофага;
- рецептор $T$-лимфоцита связывается с антигеном;
- происходит активация $T$-лимфоцита;
- в $T$-лимфоците образуется белок – интерлейкин-2;
- на мембранах $T$-клеток формируются рецепторы к интерлейкину, происходит связывание с ним;
- начинаются процессы деления и дифференцировки $T$-клеток с образование клеток-эффекторов (посредники при вовлечении в иммунный ответ $B$-лимфоцитов, $T$-хелперы) или клонов ($T$-киллеры).
$T$-лимфоциты-хелперы при участии макрофагов передают информацию об антигене $B$-клеткам. Роль макрофагов в данном процессе:
- снимают антигенрецепторные комплексы с поверхности $T$-лимфоцитов;
- концентрируют на своей поверхности эти комплексы в виде «обоймы»;
- передают $B$-лимфоциту «обоймы» антигенных молекул.
Для активации $B$-лимфоцита необходимо, чтобы молекула антигена была подготовлена макрофагом и $T$-лимфоцитом. Таким образом происходит передача специфической информации об антигене от $T$-лимфоцита $B$-лимфоциту.
В виде гуморального стимула передается второй неспецифический сигнал от $T$-клеток $B$-лимфоцитам.
Замечание 2
$B$-лимфоциты, после получения специфического и неспецифического сигналов, пролиферируют с дифференцировкой в различные клоны эффекторных клеток. При этом образуются плазматические клетки, продуцирующие иммуноглобулины.
Источник
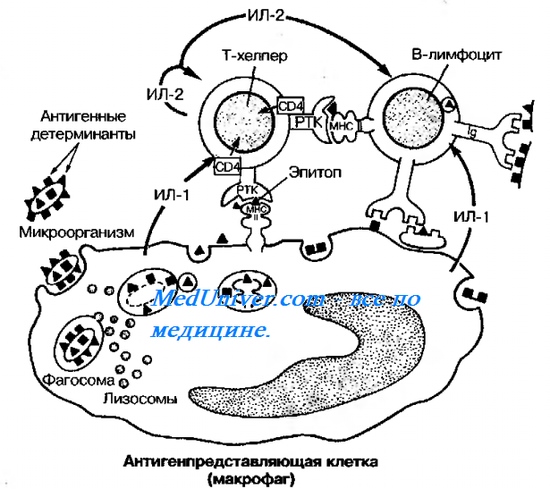
Оглавление темы “Гуморальные имунные реакции. Основные типы антител. Динамика антителообразования.”: Гуморальные имунные реакции. Антиген представляющие клетки. Т-хелпер. Взаимодействие клеток в ходе гуморального иммунного ответа. В-лимфоцит.В гуморальных иммунных реакциях участвуют три клеточных типа: макрофаги (Аг-представляющие клетки), Т-хелперы и В-лимфоциты (рис. 10-7). Антиген представляющие клеткиАг-представляющие клетки фагоцитируют микроорганизм и перерабатывают его, расщепляя на фрагменты {процессинг Аг). Фрагменты Аг выставляются на поверхности Аг-представляю-щей клетки вместе с молекулой МНС. Комплекс «Аг-молекула МНС класса II» предъявляется Т-хелперу. Распознавание комплекса Т-хелпером стимулирует секрецию ИЛ-1 макрофагами.
Т-хелперТ-хелпер под действием ИЛ-1 синтезирует ИЛ-2 и рецепторы к ИЛ-2; последний по аутокринному механизму стимулирует пролиферацию Т-хелперов, а также ЦТЛ. Таким образом, после взаимодействия с Аг-представляющей клеткой Т-хелпер приобретает способность отвечать на действие ИЛ-2 бурным размножением. Биологический смысл этого явления состоит в накоплении Т-хелперов, обеспечивающих образование в лимфоидных органах необходимого пула плазматических клеток, вырабатывающих AT к данному Аг. В-лимфоцитАктивация В-лимфоцита предполагает прямое взаимодействие Аг с молекулой Ig на поверхности В-клетки. В этом случае сам В-лимфошт перерабатывает Аг и представляет его фрагмент в связи с молекулой МНС II на своей поверхности. Этот комплекс распознаёт Т-хелпер, отобранный при помощи того же Аг. Узнавание рецептором Т-хелпера комплекса Аг-молекула МНС класса II на поверхности В-лимфоцита приводит к секреции Т-хелпером ИЛ-2, ИЛ-4, ИЛ-5 и у-ИФН, под действием которых В-клетка размножается, образуя клон плазматических клеток (плазмоцитов). Плазмоциты синтезируют AT. Секрецию AT стимулирует ИЛ-6, выделяемый активированными Т-хелперами. Часть зрелых В-лимфоцитов после антиген за виси-мой дифференцировки циркулирует в организме в виде клеток памяти. – Также рекомендуем “Антитела ( Ат ). Строение антител. Структура антител. Функционирование антител. Как устроено антитело?” |
Источник
В-лимфоциты отвечают за гуморальный адаптивный иммунный ответ, направленный преимущественно на поражение внеклеточных инфекционных агентов. При связывании специфического антигена В-лимфоциты при кооперативном взаимодействии с Т-лимфоцитами-хелперами пролиферируют, дифференцируются в плазматические клетки, секретирующие антитела, и клетки памяти. Антиген вызывает селекцию клонов В-лимфоцитов, экспрессирующих специфичные к нему BCR.
Различают В1- и В2-субпопуляции В-лимфоцитов, участвующие в реакциях врожденного и адаптивного иммунитета.
В1-лимфоциты (CD5+). Возникают в эмбриогенезе, локализуются преимущественно в брюшной и плевральной полости и lamina propria, распознают тимуснезависимые антигены и секретируют в основном IgM; не формируют клеток памяти. В отличие от «обычных» В-лимфоцитов Bl-клетки способны к самоподдержанию и играют важную роль в защите от патогенных микроорганизмов. Известны 2 субпопуляции В-1-лимфоцитов: B-la (CD5+) и B-lb (CD5).
В2-лимфоциты (CD5 ) проходят дифференцировку в эмбриональном периоде в печени, затем в костном мозге, а антигензависимый этап дифференцировки — в фолликулах периферических лимфоидных органов. В2-лимфоциты характеризуются широким разнообразием BCR, распознают Т-зависимые антигены, продуцируют иммуноглобулины разных классов, формируют иммунологическую память.
Т-лимфоциты занимают особое место в иммунной системе и служат главной популяцией в развитии клеточно-опосредованного иммунного ответа. Развитие Т-лимфоцитов зависит от тимуса, хотя выделяют также зоны внетимического развития таких клеток. Среди CD4+ и CD8+ Т-лимфоцитов выделяют следующие основные субпопуляции:
• Т-клетки-хелперы (ThO, Thl, Th2, Thl7, Tfh);
• регуляторные Т-клетки клетки (Treg, Trl, Th3);
• цитотоксические Т-клетки (ЦТЛ).
CD4+ Т-хелперы — функциональная субпопуляция Т-клеток, которые продуцируют различные цитокины и участвуют в распознавании антигенного пептида в комплексе с HLA класса II, на антигенпрезентирующей клетке, в генерации цитотоксических Т-лимфоцитов, в межклеточной кооперации с В-клетками, направляя их дифференцировку по пути плазматических клеток, синтезирующих антитела, а также в некоторых вариантах цитотоксичности.
Цитотоксические CD8+ Т-лимфоциты (ЦТЛ) — клетки-киллеры, способные поражать инфицированные вирусом клетки-мишени, опухолевые клетки, клетки трансплантата. ЦТЛ распознают антигенные пептиды в комплексе с молекулами HLA класса I.
Регуляторные CD4+ CD25+ FoxP3+ Т-лимфоциты (Treg) подразделяются на природные (естественные), развивающиеся в тимусе, и индуцированные на периферии из CD4+ ThO-клеток. Основная функция Treg-клеток — подавление (супрессия) активированных CD4+ и CD8+ Т-лимфоцитов, а также других клеток.
Т-клетки памяти (англ. memory cells) — долгоживущие лимфоциты, примированные антигеном, но не достигшие стадии терминальной дифференцировки в клетки-эффекторы. Т-клетки памяти экспрессируют молекулы CD45RO (изоформа тирозинфосфатазы) и много молекул CD44, способствующих их активной рециркуляции. При повторном контакте с тем же антигеном они отвечают намного быстрее и активнее, чем наивные лимфоциты. Клетки памяти отличаются от наивных большим сроком жизни, выраженной рециркуляцией и способностью к самоподдержанию.
В лимфатических узлах содержится основное количество Т-лимфоцитов (41,2%). Среди них CD4+ Т-клетки составляют 46,9%, CD8′ Т-лимфоциты — 35,0%. В селезенке содержится 15,2% лимфоцитов от общего количества в организме. На долю Т-хелперов приходится 6,9%, на долю ЦТЛ — 19,4%.
– Читать далее “Клетки врожденного имммунитета. Система фагоцитарных клеток”
Оглавление темы “Клетки иммунной системы”:
1. В-лимфоциты. Т-лимфоциты
2. Клетки врожденного имммунитета. Система фагоцитарных клеток
3. Нейтрофилы. Антигенпрезентирующие клетки
4. Дендритные клетки. Виды дендритных клеток
5. Различия между профессиональными антигенпредставляющими клетками. Эозинофилы
6. Тромбоциты. Органы центральной иммунной системы
7. Кровь и лимфа в иммунной системе. Тимус
8. Строение и функции тимуса. Морфология тимуса
9. Лимфатические узлы. Строение лимфатических узлов
10. Клетки лимфатических узлов. Развитие клеток лимфатических узлов
Источник
Взаимодействие Т- и В-лимфоцитов. Взаимодействие лимфоцитов и макрофаговНа основании всех изложенных данных были предложены различные гипотезы о трехклеточных системах иммунопоэза. В 1969 г. несколько авторов независимо друг от друга предположили, что процесс антителогенеза инициируется в результате кооперации трех типов клеток: клеток-предшественников (В-лимфоцитов), Т-лимфоцитов и макрофагов. При возникновении антигенного стимула В-лимфоцит-предшественник под влиянием сигналов со стороны Т-клеток-помощников и макрофагов переходит из покоящегося состояния в метаболически активную фазу, обеспечивающую клональную пролиферацию и дифференцировку в плазматическую клетку, активно синтезирующую и секретирующую антитела. Роль макрофагов существенна, по-видимому, на самом первом этапе распознавания антигена и приведения его в иммуногенную форму. Опыты с использованием антимакрофагальных сывороток показали необходимость присутствия макрофагов лишь в первые сутки после введения антигена, т. е. в момент антигенной стимуляции (Галактионов, Анфалова, 1974). Все теории индукции антителогенеза в В-лимфоцитах можно разделить на две группы: теория односигнальной и двухсигнальной активации В-лимфоцнтов. Сторонники односигнальных теорий постулируют необходимость одного сигнала для перехода покоящегося В-лимфоцита в метаболически активную фазу. Одни авторы считают, что этим сигналом является неспецифическое ми-тогенное влияние антигена, а не взаимодействие антигенных детерминантов с рецепторами на поверхности В-клетки. Иммуноглобулиновые рецепторы, по их мнению, обеспечивают лишь концентрирование антигена на определенных В-клетках, что приводит к преимущественной активации клонов, детерминированных к синтезу антител данной специфичности.
Экспериментальное обоснование этой гипотезы опирается на результаты опытов но изучению влияния тимуснезависимых антигенов на В-клетки. Большинство известных тимуснезависимых антигенов обладают свойствами поликлональных активаторов и являются митогенами для В-клеток (Doenhoff е. а., 1976). Имеются данные о том, что некоторые В-клеточные митогены вызывают поликлона льный синтез антител в отсутствие специфического антигена (Andersson е. а., 1972). Отсюда делается вывод, что сигналом, вызывающим индукцию иммунной реактивности в В-клетке, является неспецифическое митогенное воздействие, а не связывание специфических антигенных детерминант иммуноглобулиновыми рецепторами. Однако есть факты, которые не укладываются в представление о ведущей роли поликлонального воздействия в активации В-клетки. Так, существуют Т-независимые антигены, которые не вызывают митогенного эффекта или поликлонального синтеза антител, Кроме того, есть сведения об отсутствии корреляции между степенью митогенности тимуснезависимого носителя н силой иммунного ответа к присоединенному к нему гаптену. Известно, например, что пневмококковый полисахарид является лучшим митогеном, чем леван, однако ответ к динитрофенильной группе, конъюгированной с леваном, был выше, чем при конъюгировании данного гаптена с пневмококковым полисахаридом. Вторая точка зрения о природе стимулирующего сигнала заключается в том, что активация В-клеток происходит в результате взаимодействия антигенных детерминант с иммуноглооулиновыми рецепторами В-лимфоцита (Feldmann e. a., 1975 b; Klaus, Hamphrey, 1975). В случае тимуснезависимых антигенов многоточечное присоединение густо повторяющихся детерминант к поверхности В-клетки обеспечивает высокую энергию связывания антигена с В-лимфоцитом, что приводит к его активации. В случае тимуезависимых антигенов необходимая для активации структура антигенных детерминант создается с помощью Т-лимфоцитов, которые концентрируют антиген и представляют его В-клетке в поливалентной форме. Так, согласно гипотезе Фелдмана и соавторов, Т-клетки полимеризуют антиген на поверхности макрофага. Молекулы антигена связываются носителем с IgT, вырабатываемым активированными Т-лимфоцитами. Комплексы антиген — IgT присоединяются к макрофагу, в результате чего на поверхности макрофага создается обойма из антигенных молекул, ориентированных гаптенными группировками наружу. Такая структура с высокой плотностью идентичных антигенных детерминант и активирует В-клетку. При отсутствии макрофагов, являющихся местом структурной организации антигена, развивается не иммунный ответ, а толерантность. Таким образом, односигнальная модель также предусматривает кооперацию различных типов клеток при создании активирующего стимула. Существование тимуснезависимых антигенов, очевидно, не противоречит положению о необходимости взаимодействия клеток при индукции антителогенеза. – Также рекомендуем “Модель активации В-клетки. Индукция антителообразования” Оглавление темы “Супрессия иммунного ответа. Клетки супрессоры”: |
Источник