В лимфатических узлах есть т и в лимфоциты

ЦИРКУЛЯЦИЯ ЛИМФЫ
Рис. 1. Афферентные лимфатические сосуды (АЛС) доставляют лимфу с лимфоцитами из соединительных тканей или периферических лимфатических узлов в субкапсулярный синус (СС). Оттуда лимфа проходит через промежуточные синусы (ПС) в мозговые синусы (МС), чтобы попасть в эфферентные лимфатические сосуды (ЭЛС). В районе ворот (В) лимфа из субкапсулярного синуса идет прямо в мозговые синусы. Лимфа также инфильтрирует лимфоидную ткань и медленно течет от коркового (KB) к мозговому веществу (MB). Лимфоциты покидают паренхиму лимфатического узла через мозговые синусы, достигая эфферентного лимфатического сосуда. Направление циркуляции лимфы (стрелки) регулируется клапанами (К).

КРОВОСНАБЖЕНИЕ ЛИМФАТИЧЕСКОГО УЗЛА
Рис. 2. Кровоснабжение каждого лимфатического узла происходит за счет маленькой мышечной артерии (А), которая входит в лимфатический узел через его ворота (В), разветвляясь затем на артериолы (Арт), идущие через мозговые тяжи (MT). Достигнув важнейшего коркового слоя (КС), артериолы распадаются на капиллярную сеть вокруг и внутри герминативных центров лимфоидных узелков (ЛиУ). Кровь затем собирается посткапиллярными венулами (ПсКВ), которые проходят радиально через внутренний слой коркового вещества (ВКВ), входят в мозговые тяжи и объединяются в большую вену (Be), которая покидает лимфатичекий узел через его ворота. Как отмечалось ранее, посткапиллярные венулы — это места, где лимфоциты покидают поток крови, чтобы войти в паренхиму лимфатического узла.
ЛИМФОЦИТАРНАЯ ЦИРКУЛЯЦИЯ (ЦИРКУЛЯЦИЯ ЛИМФОЦИТОВ)
Рис. 3. Лимфоцитарная циркуляция — это медленное движение лимфопоэтических клеток и лимфоцитов между различными лимфоидными органами и тканями посредством крови и лимфы.
Т- и В-лимфообразующие клетки появляются из лимфоидной клетки-предшественницы (ЛКП) костного мозга. Предшественники Т-лимфоцитов (Т) мигрируют через кровоток в тимус, где дифференцируются в зрелые иммунокомпетентные Т-лимфоциты в контакте с эпителиоретикулярными клетками тимуса, находясь в особом микроокружении этого органа.
Большинство В-лимфоцитов (В), формирующихся в костном мозге, поступает в кровоток, часть клеток-предшественниц В-лимфоцитов остается в костном мозге для поддержания популяции. Отсюда они мигрируют в аналог сумки Фабрициуса* (аппендикс, пейеровы бляшки и миндалины), где и происходит их созревание. Иммунокомпетентные В-лимфоциты развиваются из этих клеток митотическим путем. Из тимуса и аналога сумки Фабрициуса Т- и В-лимфоциты опять поступают в кровоток, затем покидают его, мигрируя в ткани. Лимфоциты собираются в лимфатических узлах после прохождения через афферентные лимфатические сосуды (АЛС).
Через эфферентные лимфатические сосуды (ЭЛС) лимфоциты опять проходят в кровоток и ткани, и цикл повторяется. Лимфоцитарная циркуляция требует несколько недель.
РЕЦИРКУЛЯЦИЯ ЛИМФОЦИТОВ
Рис. 4. Лимфоцитарная рециркуляция — это явление, параллельное циркуляции лимфоцитов.
Она состоит из быстрой миграции В- и Т-лимфоцитов из крови в ткани и лимфоидные органы и затем назад в кровь.
Стрелки указывают направление лимфо- и кровотока.
В-лимфоциты (светлые кружочки) и Т-лимфоциты (темные кружочки) входят в ткани (Тk) из кровеносных капилляров (Кап), чтобы быть собранными афферентными лимфатическими сосудами (АЛС) в лимфатических узлах (ЛУ). Здесь некоторые Т-лимфоциты заселяют тимус зависимые зоны, но большинство лимфоцитов продвигается через синусы в паренхиму, поступая затем через эфферентные лимфатические сосуды (ЭЛС) в грудной проток (ГП), посредством которого они попадают в кровоток (КГ). Отсюда лимфоциты распределяются по лимфоидным органам (тимус — Т, аналог сумки—АС; селезенка —С, лимфатические узлы — ЛУ) и тканям. Т-лимфоциты образуются в тимусе, они не рециркулируют через его паренхиму, а покидают орган через его лимфатические и кровеносные сосуды. В лимфатических узлах лимфоциты проходят через стенки посткапиллярных венул (ПсКВ) и поступают в лимфоидную ткань, мигрируя через нее, чтобы вновь накопиться в эфферентных лимфатических сосудах. Лимфоцитарная рециркуляция через кровь занимает около 0,6 ч, прохождение через селезенку—около 6 ч, а через лимфатические узлы 15—20 ч. Во время этого периода рециркуляции лимфоциты не делятся. Лимфобласты не рециркулируют.
Цель лимфоцитарной рециркуляции — дать возможность иммунокомпетентным лимфоцитам осуществлять постоянную защиту организма и информировать лимфоидные органы о наличии или отсутствии антигенов в теле. В присутствии антигенов часть лимфоцитов оседает в лимфоидных органах и начинает делиться, приводя в действие иммунную реакцию. Те, которые потеряны при циркуляции, компенсируются за счет медленной лимфоцитарной циркуляции посредством воспроизводства в костном мозге, тимусе, селезенке и других лимфоидных органах.
*Сумка Фабрициуса — лимфоэпителиальный орган, обнаруженный у птиц в форме клоакального дивертикула, который участвует в производстве иммунокомпетентных В-лимфоцитов. Сумка Фабрициуса не существует у млекопитающих и человека, но предполагают, что лимфоидные узелки в миндалинах, аппендиксе, пейеровых бляшках и других органах могут частично выполнять эту роль. Поэтому эти органы называют аналогом сумки Фабрициуса, или эквивалентом сумки.
Источник
Т-лимфоциты и В-лимфоциты иммунитета. Предварительная обработка Т- и В-лимфоцитовХотя при исследовании под микроскопом большинство лимфоцитов в нормальной лимфоидной ткани выглядят одинаковыми, эти клетки подразделяют на две основные популяции. Одна популяция — Т-лимфоциты — ответственна за формирование активированных лимфоцитов, обеспечивающих клеточно-опосредованный иммунитет. Другая популяция — В-лимфоциты — ответственна за формирование антител, обеспечивающих гуморальный иммунитет. Оба типа лимфоцитов образуются у эмбриона из полипотентных гемопоэтических стволовых клеток, формирующих лимфоциты как один из наиболее важных результатов их дифференцировки. Почти все сформированные лимфоциты в результате заселяют лимфоидную ткань, однако прежде чем это произойдет, они дополнительно дифференцируются или проходят предварительную обработку. Лимфоциты, которые в итоге станут активированными Т-лимфоцитами, сначала мигрируют к тимусу, где подвергаются предварительной обработке. Эти ответственные за клеточно-опо-средованный иммунитет лимфоциты называют Т-лимфоцитами, что подчеркивает роль тимуса. Другая популяция лимфоцитов, В-лимфоциты, предназначенные для формирования антител, проходят предварительную обработку в печени плода в середине периода внутриутробной жизни, а также в костном мозге в конце внутриутробной жизни плода и после рождения. Эта популяция клеток впервые была открыта у птиц, имеющих специальный орган для их предварительной обработки, который называют бурсой Фабриция (фабрициевой сумкой). Ответственные за гуморальный иммунитет лимфоциты называют В-лимфоцитами, что подчеркивает роль бурсы. На рисунке показаны две лимфоцитарные системы для формирования: (1) активированных Т-лимфоцитов; (2) антител.
Предварительная обработка Т- и В-лимфоцитовВсе лимфоциты организма происходят от коммитированных в лимфоцитарном направлении стволовых клеток эмбриона, но эти клетки не могут непосредственно превратиться в активированные Т-лимфоциты или антитела. Прежде чем это станет возможным, клетки должны подвергнуться дальнейшей дифференцировке в соответствующих областях, где они проходят специфическую обработку. Т-лимфоциты проходят предварительную обработку в тимусе (вилочковой железе). После образования в костном мозге Т-лимфоциты сначала мигрируют к вилочковой железе. Здесь они быстро делятся, одновременно становясь чрезвычайно разнообразными, т.е. предназначенными для реакции против разных специфических антигенов. Это значит, что один лимфоцит, обработанный в тимусе, проявляет специфическую реактивность в отношении одного антигена. Следующий лимфоцит специфически реагирует на другой антиген. Это продолжается до тех пор, пока в тимусе не появятся тысячи разных типов лимфоцитов со специфической реактивностью в отношении тысяч разных антигенов. Эти разные типы предварительно обработанных Т-лимфоцитов оставляют тимус и распространяются кровью по всему телу, временно оседая в лимфоидной ткани. Кроме того, благодаря обработке в тимусе любой оставляющий его Т-лимфоцит не реагирует с белками или другими антигенами собственных тканей организма (иначе Т-лимфоциты погубили бы собственное тело человека в течение всего нескольких дней). Тимус выбирает, какие Т-лимфоциты могут его покинуть, сначала смешивая их практически со всеми специфическими аутоантигенами собственных тканей тела. Если Т-лимфоцит реагирует, он разрушается и фагоцитируется, вместо того, чтобы выделяться. Это происходит с основной частью клеток (вплоть до 90%). Таким образом, клетки, выделяющиеся из тимуса, не реагируют против собственных антигенов тела; они реагируют лишь на антигены внешних источников, например бактерий, токсинов или тканей, пересаженных от другого человека. Основная часть предобработки Т-лимфоцитов в тимусе происходит перед рождением ребенка и в течение нескольких месяцев после рождения. Удаление вилочковой железы после этого периода ослабляет (но не исключает) Т-лимфоцитарную иммунную систему. Однако удаление тимуса за несколько месяцев до рождения может нарушить развитие всего клеточно-опосредован-ного иммунитета. Поскольку именно клеточный тип иммунитета в основном отвечает за отторжение трансплантированных органов, например сердца или почек, органы можно пересаживать с меньшей вероятностью отторжения, если у животного в соответствующее время до его рождения удалить тимус. В-лимфоциты проходят предварительную обработку в печени и костном мозге. О деталях предварительной обработки В-лимфоцитов известно гораздо меньше, чем о предобработке Т-лимфоцитов. Известно, что у человека предварительная обработка В-лимфоцитов осуществляется в печени в середине внутриутробного периода развития, а также в костном мозге в конце внутриутробного периода и после рождения. Существуют два важных различия между В- и Т-лимфоцитами. Во-первых, В-лимфоциты активно секретируют реактивные агенты, называемые антителами, в отличие от Т-лимфоцитов, реагирующих с антигеном непосредственно. Антитела — это крупные белковые молекулы, способные соединяться с антигенной субстанцией и разрушать ее. Во-вторых, разнообразие В-лимфоцитов выражено больше, чем у Т-лимфоцитов, т.е. формируются миллионы типов В-лимфоцитарных антител с разными специфическими реактивностями. После предобработки В-лимфоциты, как и Т-лимфоциты, мигрируют к лимфоидной ткани по всему телу, где временно располагаются рядом, но несколько обособленно от областей локализации Т-лимфоцитов. – Также рекомендуем “Роль лимфоцитарных клонов. Происхождение клонов лимфоцитов” Оглавление темы “Клетки иммунитета. Виды иммунитета”: |
Источник
Лимфатический узел является основным органом, формирующим иммунологический ответ при проникновении чужеродных веществ в организм через кожу и эпителий слизистых оболочек. Он служит вторичным барьером на пути распространения инфекции после иммунной системы кожи и слизистых оболочек.
Структура лимфатического узла является типичным примером разобщения Т- и В-клеточных лимфоидных зон. Этот принцип во многом характерен и для селезенки, и для лимфоидной системы слизистых оболочек.
В-клетки лимфатического узла сгруппированы в компактные шаровидные образования (фолликулы), расположенные в «покоящемся» лимфатическом узле, в основном субкапсулярно. Совокупность этих В-клеточных образований локализуется в так называемой кортикальной зоне. Т-клеточная (паракортикальная) зона находится под кортикальной зоной, т. е. более удалена от капсулы лимфатического узла.
Лимфоидная ткань лимфатического узла пронизана системой синусов, в которые лимфоциты попадают с афферентной лимфой (субкапсулярный синус) и покидают узел (медуллярные синусы), поступая в эфферентные лимфатические сосуды. В лимфатическом узле представлены разные популяции фагоцитирующих (макрофаги, гистиоциты) и нефагоцитирующих (дендритные клетки) антигенпрезентирующих клеток. Они очень разнообразны и имеют тропность к Т-зонам (интердигитирующие клетки) или фолликулам лимфатического узла (фолликулярные дендритические клетки). При развитии иммунного ответа архитектоника лимфатического узла претерпевает существенные изменения.
Большинство лимфоцитов поступает в лимфатические узлы из крови через специализированный сосудистый эндотелий посткапиллярных венул (ВЭВ). Происходит это главным образом на границе кортикальной и паракортикальной зон. Другой путь поступления лимфоцитов в лимфатические узлы — через афферентные лимфатические сосуды.
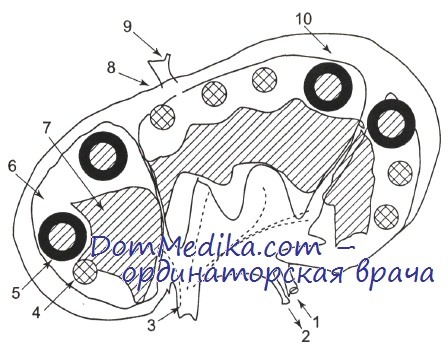
Лимфатический узел (схема).
1 — артерия; 2 — вена; 3 — выносящий лимфатический сосуд; 4 — первичный фолликул; 5 — вторичный фолликул; 6 — кортикальная зона; 7 — паракортикальная зона; 8 — капсула; 9 — приносящий лимфатический сосуд; 10 — субкапсулярный синус.
Т-лимфоциты лимфатических узлов
Наивные, т. е. еще не участвовавшие в иммунных ответах, CD4+CD8- и CD4-CD8+ Т-клетки поступают из тимуса в кровоток и через вены с высоким эндотелием проникают в лимфатические узлы.
Наивные CD4+-клетки продуцируют ИЛ-2, но практически не продуцируют ИЛ-4 или ИФу при стимуляции, обусловленной связыванием рецептора и костимуляторными сигналами. Они экспрессируют CD45RA и называются предшественниками Т-хелперных клеток. В процессе иммунного ответа наивные Т-клетки CD45RA+ (хелперные, цитотоксические) дают начало эффекторным клеткам и клеткам памяти.
В сравнении с CD45RA+ наивными Т-клетками CD45RO+ Т-клетки памяти являются функционально значительно более мощными, активируются низким содержанием антигена, не требуют костимуляторов (например, В7). CD4, CD45 и TCR более близко ассоциированы на Т-клетках памяти (CD45RO+), чем на наивных Т-клетках (CD45RA+). Подобная ассоциация делает значительно большей эффективность Т-клеточной стимуляции. Т-клетки памяти (CD45RO+) имеют сходство с активированными Т-клетками.
Это сходство заключается в экспрессии CD45RO, повышенном уровне молекул адгезии, молекул HLA II класса и рецепторов ИЛ-2. CD45RO+ Т-клетки по размеру несколько больше клеток CD45RA+, но мельче активированных Т-клеток.
Активированные Т-хелперные клетки могут дифференцироваться в ТН1 или ТН2-клетки. ТН1-клетки секретируют главным образом ИФН-у и фактор некроза опухоли-а (ФНОа). ТН2-клетки продуцируют главным образом ИЛ-4, ИЛ-5, ИЛ-6 и ИЛ-10. ТН1-клетки за счет продукции ИФН-у и ФНО являются хорошими индукторами повышения бактерицидной активности макрофагов. Эти клетки известны как клетки гиперчувствительности замедленного типа. ТН2-клоны синтезируют набор цитокинов, необходимый для гуморального иммунного ответа.
ТН2-клетки экспрессируют CD40-лиганд (CD40L), с которым связывается CD40-рецептор, присутствующий на мембране В-лимфоцитов. Связывание CD40L и действие цитокинов, секретируемых Тн2-клетками, ведет к В-клеточной пролиферации, переключению классов и развитию В-клеток памяти. Секреция ИЛ-10 и ИЛ-4 ТН2-клетками противодействует влиянию ИФН-у на макрофаги и, возможно, подавляет продукцию ИФН-у и других цитокинов ТH1-клетками. Эти отрицательные регуляторные воздействия могут быть важными в контроле аутологичного повреждения.
CD8+ Т-клетки, так же как CD4+ Т-клетки, можно подразделить на подклассы CD45RA+ и CD45RO+.
Т-клетки локализуются в паракортикальных зонах лимфатических узлов, обнаруживаются в медуллярной зоне в смеси с В-лимфоцитами и плазматическими клетками. В этих областях CD4+ Т-клетки преобладают над CD8+ Т-лимфоцитами и располагаются кластерами вокруг интердигитирующих клеток. Отдельные Т-клетки с хелперным фенотипом встречаются также в зародышевых центрах. Все лимфоциты, которые поступили в лимфатический узел из крови или лимфы, так же как клетки, образовавшиеся непосредственно в лимфатическом узле в результате клональной экспансии, покидают узел с лимфой через медуллярные синусы.
Эфферентные лимфоциты являются ответственными за становление иммунологической памяти и реализацию иммунного ответа в разных лимфоидных органах. В лимфе среди Т-клеток преобладают CD4+-лимфоциты над CD8+-лимфоцитами, что предполагает преимущественную рециркуляцию CD4+-клеток.
Иммунная активация в норме ведет к тому, что Т-клетки покидают состояние G0, входят в клеточный цикл и дают начало клону дифференцирующихся эффекторных клеток, например Т-клеток, секретирующих цитокины. Это называется клональной селекцией. Устранение аутореактивных Т-лимфоцитов называется клональной делецией. Функциональная инактивация лимфоцитов под действием антигена не обязательно сопровождается гибелью этих лимфоцитов. Состояние неотвечаемости носит название клональной анергии.
Преактивированные эффекторные Т-клетки подвергаются апоптозу, когда сталкиваются с аутологичным антигеном, представленным собственными молекулами HLA, но в отсутствие «профессиональных» антигенпрезентирующих клеток (АПК). Индукция анергии или апоптоз активированных Т-лимфоцитов также имеет место в периферических лимфоидных органах как составная часть событий посттимической негативной селекции.
Цитотоксические Т-клетки памяти экспрессируют повышенные уровни молекул адгезии, включая CD2, LFA-1, LFA-3, CD44, ICAM-1, VLA-4, VLA-5, VLA-6. Они не требуют костимуляторов В7 для активации и потому способны осуществлять киллерное действие в отношении любых презентирующих антигенный пептид клеток (например, клеток, зараженных вирусом) даже в тех случаях, если эти клетки не относятся к профессиональным АПК.
Т-лимфоциты функционально гетерогенны. Их активация приводит к Т-клеточно-опосредованным иммунным реакциям. В ходе этих реакций эффекторные Т-лимфоциты вырабатывают цитокины или осуществляют цитотоксическое действие.
– Читать “Функции Т-лимфоцитов лимфатических узлов”
Оглавление темы “Клетки крови”:
- Онтогенез Т-лимфоцитов – образование
- Созревание (онтогенез) NK-клеток
- Созревание (онтогенез) лимфоидных дендритных клеток Лангерганса
- Рециркуляция лимфоцитов в организме
- Лимфатические узлы – формирование Т-лимфоцитов
- Функции Т-лимфоцитов лимфатических узлов
- В-лимфоциты лимфатических узлов – созревание, функции
- Внутрифолликулярная дифференцировка В-лимфоцитов
- Селезенка – строение, формирование Т- и В-лимфоцитов
- Костный мозг как лимфоидный орган. Лимфопоэз в костном мозге
Источник