Транспорт липидов кровью и лимфой

Поскольку липиды не растворимы в воде, для их переноса от слизистой кишечника в органы и ткани формируются особые транспортные формы: хиломикроны (ХМ), липопротеины очень низкой плотности (ЛПОНП), липопротеины низкой плотности (ЛПНП), липопротеины высокой плотности (ЛПВП). Непосредственно от слизистой тонкого кишечника транспорт всосавшихся и ресинтезированных липидов осуществляется в составе хиломикронов. ХМ – это белково-липидные комплексы диаметром от 100 до 500 нм, которые в связи с относительно большим размером не могут сразу проникать в кровь. Сначала они попадают в лимфу и в её составе попадают в грудной лимфатический проток, а затем – в верхнюю полую вену и с кровью разносятся по всему организму. Поэтому после приёма жирной пищи плазма крови становится мутной в течение 2 – 8 часов. Химический состав ХМ: Общее содержание липидов – 97-98%; в их составе преобладают ТАГ (до 90%), на содержание холестерина (Х), его эфиров (ЭХ) и фосфолипидов (ФЛ) суммарно приходится -7-8%. Содержание белка, стабилизирующего структуру ХМ – 2-3%. Т.о., ХМ – это транспортная форма «пищевого» или экзогенного жира. В капиллярах различных органов и тканей (жировой, печени, лёгких и др.) содержится липопротеидная липаза (ЛП-липаза), расщепляющая ТАГ хиломикронов до глицерина и жирных кислот. Плазма крови при этом просветвляется, т.е. перестаёт быть мутной, поэтому ЛП-липазу называют «просветвляющим фактором». Её активирует гепарин, который вырабатывается тучными клетками соединительной ткани в ответ на гиперлипидемию. Продукты расщепления ТАГ диффундируют в адипоциты, где они депонируются или поступают в другие ткани для покрытия энергозатрат. В жировых депо по мере необходимости организма в энергии происходит распад ТАГ до глицерина и жирных кислот, которые в комплексе с альбуминами крови транспортируются к периферическим клеткам органов и тканей.
Ремнантные ХМ (т.е. оставшиеся после расщепления ТАГ) поступают в гепатоциты и используются ими для построения других транспортных форм липидов: ЛПОНП, ЛПНП, ЛПВП. Их состав дополняется жирными кислотами ТАГ, фосфолипидами, холестерином, эфирами холестерина, сфингозинсодержащими липидами, синтезированными в печени «de novo». Размер ХМ и их химический состав изменяются по мере продвижения по сосудистому руслу. ХМ обладают наименьшей по сравнению с другими липопротеинами плотностью (0,94) и наиболее крупными размерами (их диаметр ~ 100нм). Чем выше плотность ЛП-частицы, тем меньше их размер. Диаметр ЛПВП наименьший (10 – 15нм), а плотность колеблется в диапазоне 1,063 – 1,21.
ЛПОНП формируются в печени, содержат в своём составе 55% ТАГ, поэтому они считаются транспортной формой эндогенного жира. ЛПОНП транспортируют ТАГ от клеток печени к клеткам сердца, скелетных мышц, лёгких и других органов, имеющим на своей поверхности фермент ЛП – липазу.
ЛП – липаза расщепляет ТАГ ЛПОНП до глицерина и жирных кислот, превращая ЛПОНП в ЛПНП (ЛПОНП – ТАГ = ЛПНП). ЛПНП могут также синтезироваться «de novo» в гепатоцитах. В их составе преобладает холестерин (~ 50%), их функция – транспорт холестерина и фосфолипидов к периферическим клеткам органов и тканей, имеющим на своей поверхности специфические рецепторы к ЛПНП. Холестерин и фосфолипиды, транспортируемые ЛПНП, используются для построения мембранных структур периферических клеток. Поглощаясь различными клетками, ЛПНП несут информацию о содержании холестерина в крови и определяют скорость его синтеза в клетках. ЛПВП синтезируются главным образом в клетках печени. Это наиболее устойчивые формы липопротеинов, т.к. содержат ~50% белка. Они отличаются высоким содержанием фосфолипидов (~20%) и низким содержанием ТАГ (~3%). ЛПВП (см. табл. №1) синтезируются гепатоцитами в виде плоских дисков. Циркулируя в крови, они поглощают избыток холестерина от различных клеток, стенок сосудов и, возвращаясь к печени, приобретают шаровидную форму. Т.О. , основная биологическая функция ЛПВП – транспорт холестерина от периферических клеток к печени. В печени избыток холестерина превращается в желчные кислоты.
Таблица №1. Химический состав транспортных липопротеинов (%).
| Химичесий состав | Хиломикроны | ЛПОНП | ЛПНП | ЛПВП |
| Белок | 2 -3 | |||
| ТАГ | ||||
| Холестерин | 3 -5 | |||
| Фосфолипиды | 3 – 4 |
ЛП – частицы крови: ХМ, ЛПОНП, ЛПНП, ЛПВП имеют общие закономерности строения. В центре располагаются неполярные (гидрофобные ) липиды: ТАГ и эфиры холестерина образуют гидрофобное ядро. В периферическом слое располагаются полярные липиды: глицерофосфолипиды, ганглиозиды и др., образуя гидрофильный слой ЛП-частицы. Свободный холестерин своей НО-группой обращён к поверхностному гидрофильному слою частицы. Белки липопротеидных частиц принято называть апопротеинами. Различают поверхностные, периферические и интегральные белки. Последние пронизывают гидрофильный слой частицы. Различают апопротеины А, В, С, которые делятся на подгруппы. В их составе ферменты, белки – рецепторы, активаторы ферментов, апо – белки. Они определяют растворимость ЛП – частиц, их устойчивость в кровеносном русле, связь с рецепторами клеточных мембран. Виды химических связей в ЛП- частицах, чаще всего, гидрофобные и ионные. Синтез различных апопротеинов происходит в гепатоцитах, в слизистой тонкого кишечника. Толщина гидрофильного слоя – 2,2 мкМ – одинакова у всех видов ЛП-частиц. Размер частиц определяется толщиной гидрофобного ядра.
Основными апопротеинами ХМ являются – апо В 48, ЛПОНП – апо В 100, ЛПНП – апо В 100, ЛПВП – апо А. Особую роль в составе ЛПВП выполняет фермент лецитинхолестеринацилтрансфераза (ЛХАТ), катализирующий реакцию: лецитин +свободный эфир холестерина + лизолецитин
Эфиры холестерина погружаются в гидрофобное ядро ЛП-частиц, при этом в поверхностном слое освобождается место для свободного холестерина. Забирая избыток свободного холестерина от клеток и стенок сосудов, ЛП-частицы постепенно приобретают шаровидную форму и возвращаются в печень. В печени избыток холестерина превращается в желчные кислоты. Ключевым ферментом этого процесса является 7-альфа гидроксилаза.
Источник
Поскольку липиды являются в основе своей гидрофобными молекулами, то они транспортируются в водной фазе крови в составе особых частиц – липопротеинов.
Структуру транспортных липопротеинов можно сравнить с орехом, у которых имеется скорлупа и ядро. “Скорлупа” липопротеина является гидрофильной, ядро – гидрофобное.
- поверхностный гидрофильный слой формируют фосфолипиды (их полярная часть), холестерол (его ОН-группа), белки. Гидрофильность липидов поверхностного слоя призвана обеспечить растворимость липопротеиновой частицы в плазме крови,
- “ядро” формируют неполярные эфиры холестерола (ХС) и триацилглицеролы (ТАГ), которые и являются транспортируемыми жирами. Их соотношение колеблется в разных типах липопротеинов. Также к центру обращены жирнокислотные остатки фосфолипидов и циклическая часть холестерола.
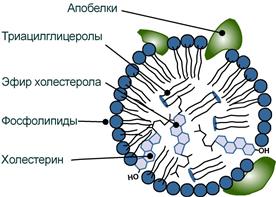
Схема строения любого транспортного липопротеина
Выделяют четыре основных класса липопротеинов:
- липопротеины высокой плотности (ЛПВП, α-липопротеины, α-ЛП),
- липопротеины низкой плотности (ЛПНП, β-липопротеины, β-ЛП),
- липопротеины очень низкой плотности (ЛПОНП, пре-β-липопротеины, пре-β-ЛП),
- хиломикроны (ХМ).
Свойства и функции липопротеинов разных классов зависят от их состава, т.е. от вида присутствующих белков и от соотношения триацилглицеролов, холестерола и его эфиров, фосфолипидов.
Подробно состав липопротеинов описан здесь
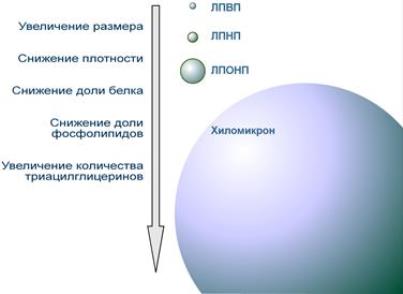
Cравнение размера и состава липопротеинов
Функции липопротеинов
Функциями липопротеинов крови являются
1. Перенос к клеткам тканей и органов
- насыщенных и мононенасыщенных жирных кислот в составе триацилглицеролов для последующего депонирования или использования в качестве энергетических субстратов,
- полиненасыщенных жирных кислот в составе эфиров холестерола для использования клетками в синтезе фосфолипидов или образования эйкозаноидов,
- холестерола в качестве мембранного материала,
- фосфолипидов в качестве мембранного материала,
Хиломикроны и ЛПОНП ответственны, в первую очередь, за транспорт жирных кислот в составе ТАГ. Липопротеины высокой и низкой плотности – за транспорт свободного холестерола и жирных кислот в составе его эфиров. ЛПВП способны также отдавать клеткам часть своей фосфолипидной оболочки.
2. Удаление избытка холестерола с мембран клеток.
3. Транспорт жирорастворимых витаминов.
4. Перенос стероидных гормонов (наряду со специфическими транспортными белками).
Апобелки липопротеинов
Белки в липопротеинах обычно называются апобелками, выделяют несколько их типов – А, В, С, D, Е. В каждом классе липопротеинов находятся соответствующие ему апобелки, выполняющие свою собственную функцию:
1. Структурная функция («стационарные» белки) – связывают липиды и формируют белок-липидные комплексы:
- апоВ-48 – присоединяет триациллицеролы,
- апоВ-100 – связывает как триацилглицеролы, так и эфиры холестерина,
- апоА-I – акцептирует фосфолипиды,
- апоА-IV – связывается с холестеролом.
2. Кофакторная функция («динамические» белки) – влияют на активность ферментов метаболизма липопротеинов в крови:
- апоС-II – кофактор гепаринзависимой липопротеинлипазы,
- апоС-III – кофактор печеночной ТАГ-липазы и ингибитор липопротеинлипазы,
- апоА-I, апоА-II и апоС-I – кофакторы лецитин-холестерол-ацилтрансферазы,
- апоЕ – ингибитор липопротеинлипазы.
3. Векторная функция (белки-маркеры, стационарные) – обеспечивают направленный транспорт липопротеинов:
- апоВ-48, апоВ-100 и апоА-I – связываются со своими рецепторами клеток-мишеней,
- апоЕ помогает взаимодействию векторных апобелков с рецепторами.
Названия апобелков B-48 и B-100 возникли не случайно. Длина мРНК гена апоВ равна 14 тысяч оснований и молекулярная масса “целого 100%-го” белка апоB-100 составляет 512 кДа. В мРНК имеется глутаминовый кодон СAA, который в результате процессинга мРНК превращается в терминирующий кодон UAA (стоп-кодон). В результате трансляции этой мРНК образуется укороченный апоB – апоВ-48 (241 кДа), который содержит N-концевые аминокислотные остатки апоВ-100.
Источник
Жиры гидрофобны, поэтому существуют специальные механизмы их транспорта в крови. Свободные (неэстерифицированные) жирные кислоты переносятся кровью в виде комплексов с альбуминами. Холестерол, его эфиры, триацилглицеролы, фосфолипиды транспортируются в составе липопротеинов.
Липопротеины являются молекулярными комплексами, состоящими из липидов и белков.
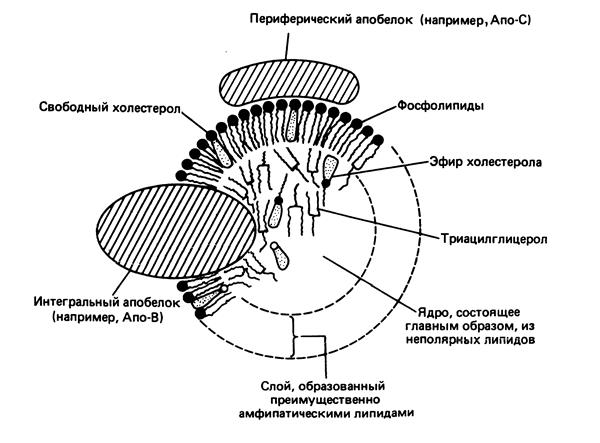
Рис. 10.2. Строение липопротеина
Существует несколько классов липопротеинов (ЛП), но всех их объединяют следующие особенности: 1) поверхностный слой липопротеинов состоит из фосфолипидов, свободного холестерола и белков; 2) каждый липопротеин содержит особый набор поверхностных белков – аполипопротеинов (апо), которые обозначаются буквами латинского алфавита (А,В,С); 3) сердцевина (ядро) липопротеина состоит из гидрофобных триацилглицеролов, эфиров холестерола (рис. 10.2 ).
Аполипопротеины выполняют следующие функции: 1) являются структурными компонентами липопротеинов; 2) участвуют в узнавании и взаимодействии с рецепторами мембран; 3) активируют ферменты метаболизма липопротеинов.
Липопротеины подразделяются на 4 основные класса в зависимости от плотности (определяемой с помощью ультрацентрифугирования) и электрофоретической подвижности (табл. 10.1).
Таблица 10.1.
Классификация липопротеинов по методу разделения
| Класс | Ультрацентрифугирование | Электрофорез |
| Хиломикроны | Хиломикроны | Хиломикроны |
| Липопротеины очень низкой плотности | ЛПОНП (VLDL) | пре-b-ЛП |
| Липопротеины низкой плотности | ЛПНП (LDL) | b-ЛП |
| Липопротеины высокой плотности | ЛПВП (HDL) | a-ЛП |
Основные параметры и состав липопротеинов представлен в табл. 10.2.
Хиломикроны (ХМ) – самые крупные частицы. ХМ синтезируются в слизистой кишечника и участвуют в экзогенном транспорте пищевых липидов к различным тканям. Основным липидом являются триацилглицеролы.
ЛПОНПсинтезируются в печени. Основным липидом являются триацилглицеролы. Основная функция – транспорт эндогенных липидов из печени в периферические ткани.
ЛПНП образуются в кровеносном русле из ЛПОНП. Содержат много холестерола (основной транспортер холестерола), который транспортируется в периферические ткани.
ЛПВП образуются в печени, содержат много фосфолипидов и белков; у этих ЛП компоненты оболочки преобладают над сердцевиной.
Таблица 10.2
Состав липопротеинов
| Липопротеины | Молекулярная массы, Да | Диаметр, нм | Состав, % | |||
| белок | ТГ | ФЛ | ХС | |||
| Хиломикроны | 1-10 млн. | 100-1000 | 1-2 | 88-90 | 4-7 | 5-6 |
| ЛПОНП | 5-100 тыс. | 30-90 | 7-10 | 50-56 | 20-23 | |
| ЛПНП | 2-4 млн. | 20-25 | 20-21 | 10-13 | 21-24 | 45-47 |
| ЛПВП | 200-400 тыс. | 10-15 | 35-50 | 5-8 | 30-43 | 20-35 |
ТГ – триацилглицеролы, ФЛ – фосфолипиды. ХС – холестерол
Различают экзогенный (транспорт пищевых липидов) и эндогенный (транспорт липидов, синтезированных в организме) транспорт.
Экзогенный транспорт. Продукты переваривания липидов всасываются в клетки слизистой оболочки кишечника в составе мицелл. Жирные кислоты с числом углеродных атомов <12 всасываются в кровь и по воротной вене транспортируются в печень. Длинноцепочечные жирные кислоты (С >12) в клетках кишечника реэстерифицируются в триацилглицеролы, которые по составу напоминают пищевые жиры. Полученные триацилглицеролы вместе с фосфолипидами, холестеролом и белками (2%) образуют хиломикроны. Хиломикроны содержат апопротеин В48 и апоА.
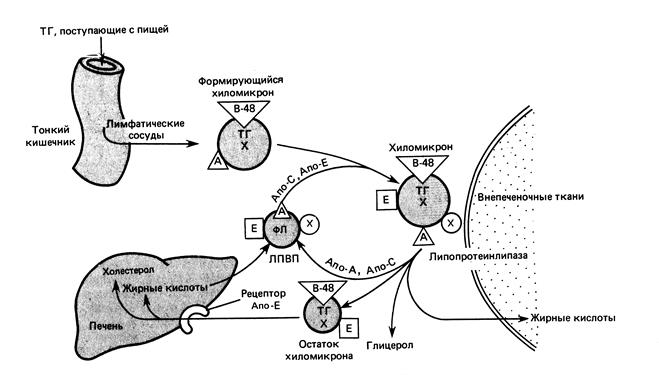
Рис. 10.3. Экзогенный транспорт липидов (по Марри Р. и др., 2004)
Хиломикроны поступают в лимфу. В крови встречаются с частицами ЛПВП, содержащими апоЕ и апоС. Хиломикроны отдают апоА частицам ЛПВП, а взамен приобретают апоЕ и апоС. Один из аполипопротеинов группы С – апоСII – служит активатором фермента липопротеинлипазы (ЛПЛ). Этот фермент синтезируется и секретируется жировой и мышечной тканями, клетками молочных желез. Секретируемый фермент прикрепляется к плазматической мембране эндотелиальных клеток капилляров тех тканей, где он синтезировался. АпоСII, находящийся на поверхности ХМ, активирует ЛПЛ. Она гидролизирует триацилглицеролы в составе ХМ до глицерола и жирных кислот. Эти жирные кислоты либо поступают в клетки жировой и мышечной ткани, либо соединяются с альбуминами плазмы. В результате действия ЛПЛ хиломикроны резко уменьшаются в размерах и их называют ремнанты (остаток). Ремнанты ХМ рецепторным путем захватываются печенью (рис. 10.3).
Эндогенный транспорт. В клетках печени ресинтезируются триацилглицеролы и фосфолипиды, которые характерны для данного организма. Они включаются в состав ЛПОНП. В состав ЛПОНП входят апоВ100 и апоС. Это основная транспортная форма триацилглицеролов. В другой класс липопротеинов, образуемых в печени – ЛПВП входят холестерол, фосфолипиды, апоА. Эти частицы плоские и их называют – насцентные ЛПВП. (В их ядре нет гидрофобных молекул). Эти ЛПВП играют большую роль в обратном транспорте холестерола из клеток периферических тканей в печень.
В капиллярах жировой и мышечной тканей апоСII ЛПОНП активирует ЛПЛ, которая катализирует гидролиз триацилглицеролов ЛПОНП и превращает их в ЛППП (липопротеины промежуточной плотности). ЛППП под действием синтезированной в печени циркулирующей печеночной триацилглицероллипазы, теряют еще часть триацилглицеролов и превращаются в ЛПНП. Основным липидом ЛПНП становится холестерол, который в составе ЛПНП переносится к клеткам всех тканей. Следовательно, ЛПНП образуются непосредственно в сосудистом русле (рис. 10.4).
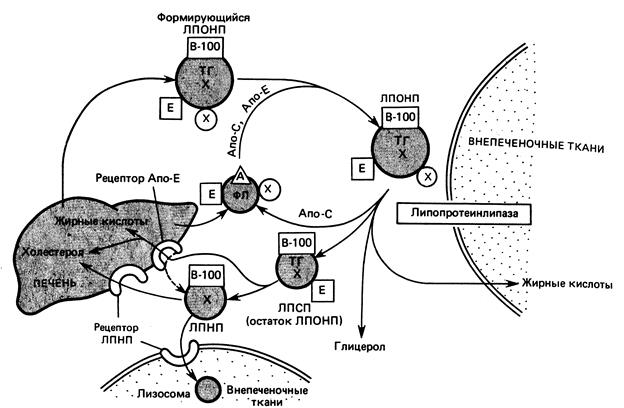
Рис. 10.4. Эндогенный транспорт липидов (по Марри Р. и др., 2004)
Итак, в результате экзогенного и эндогенного транспорта в капиллярах жировой и мышечной тканей освобождаются жирные кислоты и глицерол. Жирные кислоты связываются с альбуминами и транспортируются к тканям-потребителям.
Источник
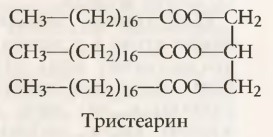
Метаболизм жиров в организме. Транспорт липидовНекоторые химические вещества, входящие в состав пищи и тканей тела, классифицируют как липиды. К ним относят: (1) нейтральные жиры, известные как триглицериды; (2) фосфолипиды; (3) холестерол; (4) некоторые другие вещества, менее важные. Основной частью химической структуры триглицеридов и фосфолипидов являются жирные кислоты, представляющие собой простые углеводородные органические кислоты с длинной цепочкой. Так, типичная жирная кислота — пальмитиновая, она может быть представлена как СН3(СН2)14СООН. Холестерол не содержит жирных кислот, но его стерольное ядро образовано частью молекулы жирной кислоты, что обусловливает его физические и химические свойства, характерные для вещества, относящегося к липидам. Организм использует триглицериды главным образом в качестве источника энергии для различных метаболических процессов, что функционально роднит их с углеводами. Однако некоторые липиды, особенно холестерол, фосфолипиды и небольшая часть триглицеридов, используются организмом в формировании мембран и прочих структурных компонентов клеток, т.е. выполняют пластические функции. а) Основа химического строения триглицеридов (нейтральных жиров). Поскольку в данных статьях по физиологии на сайте по большей части рассматриваются вопросы, связанные с использованием триглицеридов в качестве источника энергии, необходимо создать представление о химической структуре этих веществ.
Обратите внимание, что 3 молекулы жирных кислот с длинной цепочкой связаны с 1 молекулой глицерола, образуя типичную структуру триглицерида. В образовании триглицеридов в организме человека чаще всего участвуют три жирные кислоты: (1) стеариновая кислота (см. формулу тристеарина), которая включает цепочку из 18 углеродных фрагментов с полностью насыщенными водородом связями; (2) олеиновая кислота, также состоящая из 18-углеродной цепочки, но имеющей одну двойную связь в середине цепочки; (3) пальмитиновая кислота, включающая 16 атомов углерода с полностью насыщенными связями. б) Транспорт липидов в жидких средах организма. Транспорт триглицеридов и других липидов из желудочно-кишечного тракта посредством лимфы. Хиломикроны. Почти все жиры, присутствующие в пище, за исключением жиров, содержащих жирные кислоты с короткой цепочкой, всасываются из кишечника в лимфу. Во время пищеварения большинство триглицеридов расщепляются до моноглицеридов и жирных кислот. Затем во время прохождения через эпителиоциты кишечника моноглицериды и жирные кислоты ресинтезируются в новые молекулы триглицеридов, которые попадают в лимфу в виде мелкодисперсных капелек, названных хиломикронами. Диаметр хиломикронов колеблется от 0,08 до 0,6 мкм. Небольшие количества апопротеина В абсорбируются на наружной поверхности хиломикронов. Часть молекулы белка, оставшаяся свободной, выступает в водную фазу, что увеличивает суспензионную стабильность хиломикронов в лимфе и препятствует их прилипанию к стенкам лимфатических сосудов. Большая часть холестерола и фосфолипидов, всасываемых из желудочно-кишечного тракта, входит в состав хиломикронов. Таким образом, хиломикроны состоят главным образом из триглицеридов, а также содержат 9% фосфолипидов, 3% холестерола и около 1% апопротеина В. Образующиеся хиломикроны затем транспортируются вверх по грудному протоку и вместе с лимфой попадают в кровеносную систему в области впадения яремной и подключичной вен.
в) Извлечение хиломикронов из крови. Почти через час после приема пищи, содержащей большое количество жира, концентрация хиломикронов в плазме может увеличиться и составить от 1 до 2% общего количества плазмы. Из-за больших размеров хиломикронов плазма становится мутной и иногда желтой, но поскольку период полураспада хиломикронов составляет меньше 1 ч, плазма вновь становится прозрачной через несколько часов. Жиры, содержащиеся в хиломикронах, извлекаются следующим образом. 1. Триглицериды хиломикронов гидролизуются липопротеинлипазой. Жиры хранятся в клетках жировой ткани и клетках печени. Большая часть хиломикронов извлекается из циркулирующей крови во время прохождения по капиллярам жировой ткани или печени. Как жировая ткань, так и печень содержат большое количество фермента липопротеинлипазы. Этот фермент особенно активен в эндотелии капилляров, где он гидролизует триглицериды хиломикронов, когда те контактируют с эндотелием капиллярной стенки, что приводит к высвобождению жирных кислот и глицерола. Жирные кислоты, обладая способностью проникать через мембраны клеток, легко диффундируют через мембраны адипоцитов жировой ткани в клетки печени. Оказавшись внутри клеток, жирные кислоты вновь превращаются в триглицериды, взаимодействуя с глицеролом, образующимся в результате метаболических процессов в клетках, выполняющих функции депонирования (что будет рассмотрено далее). Липопротеин-липаза вызывает также гидролиз фосфолипидов, что, в свою очередь, приводит к выделению жирных кислот, преобразующихся в триглицериды и депонирующихся, как уже обсуждалось. – Также рекомендуем “Свободные жирные кислоты. Транспорт свободных жирных кислот” Оглавление темы “Синтез АТФ. Обмен глюкозы и жиров”: |
Источник