Т супрессорная функция лимфоцитов

Регуляторные Т-лимфоциты, Т-регуляторные клетки, Т-супрессоры (англ. regulatory T cells, suppressor T cells, Treg) — центральные регуляторы иммунного ответа. Основная их функция — контролировать силу и продолжительность иммунного ответа через регуляцию функции Т-эффекторных клеток (Т-хелперов и Т-киллеров).
Эти Т-лимфоциты экспрессируют FOXP3 — транскрипционный фактор, регулирующий транскрипцию генов, ответственных за дифференцировку Т-клеток и экспрессию цитокинов и других факторов, участвующих в супрессии иммунного ответа. Поэтому эти клетки часто называют регуляторными Т-клетками FOXP3+ (англ. FOXP3+ Treg cells). Кроме того, важным маркером Т-регуляторных клеток является экспрессия на их поверхности рецептора к цитокину IL-2 — CD25, соответственно их обозначают как CD25+-клетки. Помимо этих основных маркеров, клетки Treg на своей мембране экспрессируют CD62L, различные изоформы мембрано-связанной фосфатазы CD45. Различают несколько разных типов регуляторных Т-клеток: естественные Т-регуляторные клетки (T-reg1) и индуцибельные Т-регуляторные клетки (iT-reg). Индуцибельные Т-регуляторные клетки образуются под влиянием различных факторов на периферии, например, в региональных лимфатических узлах.
История открытия и феномен иммуносупрессии[править | править код]
Феномен супрессии иммунного ответа был известен очень давно, и важно было выявить его механизмы, поэтому было предположено существование специфических клеток Т-супрессоров. Однако существование этих клеток долгое время не было подтверждено экспериментально. Лишь в конце 1990-х и в начале 2000-х годов было показано существование определенных Т-клеток, которые характеризовались фенотипом CD25+FOXP3+ и эффективно подавляли иммунный ответ, как это было изначально показано на моделях аутоиммунных заболеваний.
Механизмы иммуносупрессии[править | править код]
Для супрессии иммунного ответа клетки Тreg выделяют цитокины: TGF-beta, IL-10, IFNγ, IL-35, а также экспрессируют на своей поверхности рецептор CTLA-4. Существуют несколько механизмов супрессии: прямой, при непосредственном контакте между клетками, и дистантный, осуществляющийся на расстоянии — например, через растворимые цитокины. Мишенями действия Тreg-клеток являются как Т-эффекторные клетки, так и дендритные клетки, ответственные за презентацию антигена и активацию Т-клеток. При дистантном механизме цитокины, выделяемые Тreg-клетками, например, трансформирующий ростовой фактор бета (TGF-beta), связываются со своими рецепторами на поверхности Т-эффекторных клеток и ингибирует их активацию, тем самым супрессируя иммунный ответ. Важным механизмом также служит захват IL-2 при помощи CD25 — рецептора к IL-2 и секвестрация рецептора у эффекторных Т-клеток, что препятствует активации после связывания комплекса MHC с антигеном, так как известно, что IL-2 является основным аутокринным стимулирующим фактором, поддерживающим дифференцировку и клональную экспансию Т-клеток.
Тreg-клетки, взаимодействуя с рецептором CD86 на дендритных клетках при помощи CTLA-4, способны ингибировать функцию активации дендритными клетками Т-клеток.
При прямом механизме супрессии Treg взаимодействуют с эффекторными Т-клетками и гранзим B действует через перфорины, образующие канал, вызывая апоптоз в этих клетках, тем самым элиминируя активные Т-клетки.
Литература[править | править код]
- Tang Q, Bluestone JA. The Foxp3+ regulatory T cell: a jack of all trades, master of regulation. Nat Immunol. 2008 Mar;9(3):239-244. doi:10.1038/ni1572. PMID 18285775.
Источник
Оглавление темы “СD8 лимфоциты. Антиген ( Аг ) представляющие клетки. Классификация антигенов ( Аг ).”:
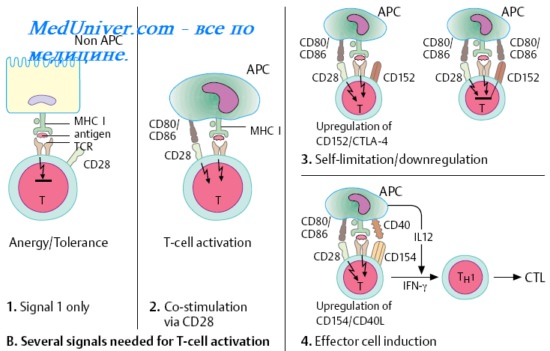
1. Рецептор Т-клеток. Активация Т-лимфоцитов. Субпопуляции Т-лимфоцитов. СD4+ лимфоциты.
2. СD8 лимфоциты. Т супрессоры. Цитотоксические Т-лимфоциты ( ЦТЛ ). Т-киллеры. Цитотоксический эффект Т-киллеров. Т-клетки памяти.
3. В-лимфоциты. В-клетки. Созревание В-лимфоцитов. Продолжительность жизни B-лимфоцитов. Маркеры В-клеток ( В-лимфоцитов ).
4. Естественные киллеры ( NK-клетки ). Перфорины. Функция естественных киллеров. Задачи NK-клеток.
5. Антиген ( Аг ) представляющие клетки. Иммунные реакции. Классификация имунных реакций. Антигены. Свойства антигенов. Структура антигенов ( Аг ). Валентность Аг.
6. Классификация антигенов ( Аг ). Иммуногены. Гаптены. Непреципитирующие гаптены. Преципитирующие гаптены. Полугаптены. Проантигены.
7. Адъюванты. Адъювант Фройнда. Толерогены. Природа антигенов (Аг). Происхождение антигенов. Молекулярная масса антигенов (Аг).
8. Специфичность антигенов (Аг). Видовые антигены. Групповые антигены (Аг). Гетерогенные антигены (Аг). Антиген Форссмана. Аллоантигены ( изоантигены ).
9. Патологические антигены. Аутоантигены. Врождённые аутоантигены. Приобретённые аутоантигены. Суперантигены.
10. Антигены ( Аг ) микроорганизмов. Антигены бактерий. Капсульные антигены ( К-антигены ( Аг )). Соматические антигены ( O-антигены ( Аг )). Жгутиковые антигены ( H-антигены ( Аг )). Vi-Ar ( Антиген вирулентности ).
СD8 лимфоциты. Т супрессоры. Цитотоксические Т-лимфоциты ( ЦТЛ ). Т-киллеры. Цитотоксический эффект Т-киллеров. Т-клетки памяти.
СD8+-лимфоциты. Мембранные Аг CD8 экспрессируют субпопуляции Т-клеток, разделяемые на регуляторные (супрессоры) и эффекторные (цитотоксические Т-лимфоциты).
Т супрессоры.
Т-супрессоры [от англ. to supress, подавлять) регулируют интенсивность иммунного ответа, подавляя активность СD4+-клеток. Т-супрессоры предотвращают развитие аутоиммунных реакций, защищают организм от нежелательных последствий иммунных реакций. Эти клетки обеспечивают толерантность матери к чужеродным Аг, представленным на клетках вынашиваемого плода. Это даёт возможность развиваться чужеродному в иммунном отношении плоду в организме матери. Т-супрессоры дифференцируются из предшественников в результате антигенной стимуляции. По-видимому, активация супрессоров находится вне контроля МНС, и представления Аг макрофагом не требуется. Аутоантигены могут стимулировать развитие ауторегуляторных Т-супрессоров.
Чужеродные Аг в неиммуногенной форме (гаптены) или иммуногенные Аг в очень высокой концентрации также способны индуцировать специфическую супрессорную активность клеток. После распознавания Аг зрелые лимфоциты препятствуют развитию иммунного ответа, действуя непосредственно на клетки или секретируя супрессорные факторы.
Цитотоксические Т-лимфоциты ( ЦТЛ ). Т-киллеры. Цитотоксический эффект Т-киллеров.
Цитотоксические Т-лимфоциты (ЦТЛ), или Т-киллеры [от англ. to kill, убивать] лизируют клетки-мишени, несущие чужеродные или видоизменённые аутоантигены (например, клетки опухолей, трансплантатов, инфицированные вирусами, клетки, несущие поверхностные вирусные Аг). В большинстве случаев функция ЦТЛ также МНС-рестригирована — цитотоксические Т-лимфоциты распознаёт чужеродный вирусный, опухолевый или трансплантационный Аг в комплексе с молекулой МНС I на мембране клетки-мишени. Индукция цитотоксических свойств клетки-предшественницы Т-киллера происходит под действием двух сигналов.
Первый сигнал включает взаимодействие между двумя комплексами: поверхностной молекулой CD8 лимфоцита и комплексом эпитоп-молекула МНС I на клетке-мишени. Второй сигнал — ИЛ, секретируемые близлежащими макрофагами и Т-клетками. Т-хелпер играет ключевую роль в стимуляции цитотоксических Т-лимфоцитов в качестве источника необходимых цитокинов, усиливающих их пролиферацию и созревание до функционально активных ЦТЛ.
Цитотоксический эффект Т-киллеров реализуется через образование в клетках-мишенях пор под действием особых белков — перфоринов. Нарушение осмотического баланса с внеклеточной средой приводит к гибели клетки.
Т-клетки памяти
Т-клетки памяти образуются при первичном иммунном ответе. Специфически распознают Аг и участвуют в иммунном ответе при вторичном попадании Аг. Большинство клеток памяти обладает функциями Т-клеток, экспрессируют CD4 и рестригированы по молекулам МНС II, то есть узнают Аг только на Аг-представляющих клетках в связи с молекулой МНС II.
– Также рекомендуем “В-лимфоциты. В-клетки. Созревание В-лимфоцитов. Продолжительность жизни B-лимфоцитов. Маркеры В-клеток ( В-лимфоцитов ).”
Источник
СD8+Т-лимфоциты — клетки, экспрессирующие антиген CD8, представлены основными двумя субпопуляциями — цитотоксическими Т-клетками и Т-лимфоцитами с супрессорной активностью.
Со временем стало известно, что CD8 экспрессируют не только эти субпопуляции лимфоцитов, но и отдельные клоны других клеток: макрофаги, естественные киллеры (ЕК), тучные клетки, дендритные клетки (ДК); основные лиганды CD8 — а2- и а3-домены антигенов I класса главного комплекса гистосовместимости (ГКГ).
Из этого следует, что CD8 — неспецифический маркер цитотоксических лимфоцитов (ЦТЛ) и Т-супрессорных лимфоцитов, но рассматривается как один из основных фенотипических признаков этих клеток.
Именно поэтому при объективной оценке уровня супрессии Т-лимфоцитов необходимо обязательно изучать супрессорную активность клеток, экспрессируюших CD8, с использованием разработанного для этой цели метода, так как только определение CD8 не дает оснований говорить ни о цитотоксической, ни о супрессорной активности Т-лимфоцитов, имеющих общий маркер CD8.
Общие представления о СD8+СD28-Т-лимфоцитах
Общее представление о супрессорных Т-лимфоцитах начало формироваться уже в 70-х годах прошлого столетия и к середине 80-х годов стало известно, что эти клетки представлены различными клонами, отличающимися условиями возникновения, кинетикой формирования, особенностями действия, разнообразием свойств, секретируемыми медиаторами и др.
Тем не менее уже тогда Б.Д. Брондзом было сформулировано, что они имеют общие особенности, которые состоят в способности блокировать дифференцировку и активность других лимфоидных клеток и это принципиально отличает их от ЦТЛ. К отличиям Т-супрессоров от других Т-лимфоцитов следует также отнести их нестабильность, высокую чувствительность к различным воздействиям, короткий период жизни и др.
Благодаря исследованиям многих известных иммунологов того времени были идентифицированы (с помощью методических возможностей того периода) некоторые поверхностные маркеры этих клеток, установлены их отличия от других клеток, выявлены некоторые этапы и механизмы активации Т-супрессоров.
В результате было сделано заключение, что Т-супрессоры и их различные клоны представляют собой регуляторные клетки, которые и осуществляют контроль соотношения между клеточным и гуморальным иммунитетом и во многом определяют интенсивность ответа к опухолям, трансплантатам, вирусам. К этому следует добавить, что указанное представление не претерпело принципиальных изменений и по настоящее время.
Со временем изучение СD8+Т-лимфоцитов дало возможность установить, что для СD8+Т-лимфоцитов с супрессорной активностью характерно отсутствие экспрессии молекул CD28, поэтому их фенотип был определен как CD8+CD28-.
При исследовании этих клеток в различных системах (особенно часто использовали смешанную культуру лимфоцитов) было показано, что они обладают множеством ингибирующих эффектов: ингибиция пролиферации СD4+Т-лимфоцитов, стимулированных аллогенными клетками, ингибиция рецепторов, связанных преимущественно с активацией клеток (рецепторы IL-2 и трансферина), подавление экспрессии ко-стимулирующих молекул антигенпрезентирующими клетками, что препятствует их оптимальному взаимодействию с СD4+Т-лимфоцитами, неспособность поддерживать секрецию цитокинов и др.
Было подтверждено, что при осуществлении своих ингибиторных влияний СD8+СD28-Т-лимфоциты распознают комплекс антигены I класса ГКГ — пептиды с участием TCR этих клеток. Установлено также, что СD8+СD28-Т-лимфоциты — гетерогенная субпопуляция.
В общей характеристике клеток этого типа важное значение имеет то, что они характеризуются снижением пролиферации в ответ на стимулы, ингибируют цитотоксичность, отличаются высоким уровнем экспрессии CD11b, CD29, CD57, CD94 при низком по сравнению с СD8+СD28+Т-лимфоцитами уровне CD25; у СD8+СD28-Т-лимфоцитов периферической крови существенно снижено фосфорилирование TCR-zeta-цепи и высокий уровень ингибитора циклинзависимой киназы р16.
Получение моноклональных антител, которые строго взаимодействуют с СD8+СD28-Т-лимфоцитами, позволило подтвердить, что они представляют собой самостоятельный клон клеток, отличающийся от цитотоксических лимфоцитов; применение указанных антител обрывало ингибирующие эффекты CD8+CD28-T-лимфоцитов как in vivo, так и in vitro, но не влияло на функции ЦТЛ.
С помощью моноклональных антител в этих клетках был идентифицирован один из эпитопов ганглиозидов — CD75s, который не выявлялся на других клетках, что послужило основанием для расширения фенотипической характеристики, которая была определена как CD8+CD28-CD75s+.
С позиций понимания общебиологического значения CD8+CD28-Т-лимфоцитов важна их способность взаимодействовать с эпителиальными клетками слизистой оболочки. CD8+CD28-T-клетки с такой способностью экспрессируют CD101 и CD103, взаимодействуют с эпителиальными клетками через белок р180 и выполняют регуляторные функции.
Авторы с полным основанием заключают, что в слизистой оболочке есть СD8+СD28-СD101+СD103+Т-лимфоциты, которые осуществляют локальный иммунологический контроль. Из этих данных следует, что регуляторные влияния СD8+СВ28-Т-лимфоцитов не ограничиваются Тh1-лимфоцитами и имеют более широкий спектр действия.
Взаимодействие СD8+СD28-Т-лимфоцитов с эпителиальными клетками представлено на рис. 58.
Рис. 58. Взаимодействие супрессорных Т-лимфоцитов и эпителиальных клеток:
CD101 — гликопротеин, участвующий в ко-стимуляции; CD103 — антиген лимфоцитов слизистой оболочки
Эти общие представления о СD8+СD28-Т-лимфоцитах в последнее время пополняются новыми данными, которые раскрывают как ранее не известные их ингибиторные эффекты, так и некоторые пути и механизмы их реализации. Такие данные получены и в экспериментальных исследованиях, и при изучении супрессорных Т-лимфоцитов периферической крови здоровых лиц, а также при некоторой патологии.
Значительный интерес представляют сведения о том, что упомянутый выше факт гетерогенности этого клона клеток получает новое освещение. При исследовании СD8+СD28-Т-лимфоцитов периферической крови человека были выявлены три их типа, объединенные способностью ингибировать антигенспецифический ответ Т-лимфоцитов.
Первый тип характеризуется способностью повреждать экспрессию ко-стимулирующих молекул на ДК — эффект, который требует непосредственного межклеточного взаимодействия. Второй обладает выраженной способностью ингибировать секрецию таких цитокинов, как IFNy и IL-6, что происходит без обязательного межклеточного контакта. Третий опосредует свои эффекты, секретируя IL-10.
При изучении как экспрессирующих CD28, так и неэкспрессирующих Т-лимфоцитов в периферической крови людей различных возрастных групп были получены важные данные. Во-первых, показано, что с возрастом количество СD8+СD28-Т-лимфоцитов уменьшается, во-вторых, стимуляция фитогемагглютинина (ФГА) увеличивает соотношение этих клонов клеток во всех возрастных группах и усиливает пролиферацию не только CD8+CD28+-, но CD8+CD28-T-клеток при более высоком уровне пролиферации последних у пожилых лиц.
Наконец, было установлено, что обработка лимфоцитов ФГА приводит к апоптозу всех СD8+Т-лимфоцитов и количество погибших клеток было одинаковым в обоих клонах — свидетельство того, что способность к апоптозу не зависела от возраста. К этому следует добавить данные о том, что увеличением количества СD8+СD28-Т-лимфоцитов у пожилых лиц без выявленной патологии можно объяснить снижение пролиферации, сопровождающееся усилением активности циклинзависимой протеинкиназы р16.
Вполне обоснованно полагать, что эти новые данные при дальнейшем изучении возрастных изменений Т-супрессорных лимфоцитов могут оказаться существенными для выяснения их роли в возрастных особенностях регуляции иммунологического гомеостаза.
К настоящему времени стали известны основные этапы ингибирующего действия СD8+СD28-Т-лифоцитами, активация которых может происходить под влиянием аллогенных, ксеногенных, а также гетерогенных антигенпрезентирующих клеток, нагруженных антигенами. Основными участниками реализации ингибирующего действия супрессорных Т-лимфоцитов являются: СD8+СD28+Т-лимфоциты, антигенпрезентирующие клетки и СD4+Т-хелперы.
При этом антигенпрезентируюшие клетки выполняют роль своеобразного мостика между Т-супрессорами и СD4+Т-лимфоцитами. Общий механизм ингибирующего действия супрессорных Т-лимфоцитов можно представить так: активация СD8+СD28-Т-лимфоцитов человека в результате распознавания ими комплекса антигены главного комплекса гистосовместимости — пептид (процесс происходит с участием TCR супрессорных клеток) на антигенпрезентирующих клетках, что лишает их способности экспрессировать ко-стимулирующие молекулы и поэтому после распознавания Т-лимфоцитами-хелперами комплекса антигены II класса ГКГ — пептид они не получают необходимого ко-стимулирующего сигнала, становятся энергичными и не способными к активации и пролиферации. Схематически этот процесс представлен на рис. 59.

Рис. 59. Этапы ингибирующего влияния СD8+СD28~Т-лимфоцитов на СD4+Т-лимфоциты (хелперы): АПК — антигенпрезентирующая клетка
Для понимания механизма действия супрессорных Т-лимфоцитов важно также то, что для осуществления ингибирующего действия они не нуждаются ни в пролиферации, ни в синтезе белка. При реализации ингибирующих сигналов Т-супрессорных лимфоцитов в антигенпрезентирующих клетках ингибируется активность NF-kappaB, что играет главную роль в их неспособности посылать ко-стимулирующие сигналы Th-лимфоцитам.
Как уже отмечено, существенное внимание уделяется вопросу о том, необходим ли для осуществления ингибирующего влияния CD8+CD28-T-лимфоцитами их непосредственный контакт с антигенпрезентирующими клетками. В настоящее время большинство авторов склоняется к тому, что такой межклеточный контакт необходим.
Множество фактов, позволяющих расширить представления о регуляторных возможностях СD8+СD28-Т-лимфоцитов, получены при их изучении в условиях трансплантации ткани, а также аутоиммунной патологии. Согласно полученным данным наличие CD8+CD28-Т-лимфоцитов во многом предопределяет судьбу трансплантата.
Сравнительное изучение этих клеток периферической крови здоровых лиц и людей с трансплантированным сердцем показало значительно более выраженную их активацию у больных, чем у здоровых лиц с параллельной активной экспрессией CD38, лейкоцитарного антигена DR, более высоким содержанием перфоринположительных клеток.
Отмечено, что экспрессия CD27 более выражена на клетках CD8+CD28- больных, у которых не происходило отторжения трансплантата, по сравнению с больными, у которых наблюдались признаки отторжения. В связи с этими данными определяется еще один аспект значения Т-супрессорных клеток: существует клон регуляторных клеток CD8+CD28-CD27+, которые играют роль в защите трансплантата.
Установлено также, что эти клетки, выделенные из трансплантатов, не проявляют цитотоксической активности против клеток донора и имеют более высокий уровень экспрессии KIR94 — факты, свидетельствующие о том, что при трансплантации происходят фенотипические изменения в Т-супрессорных лимфоцитах.
Учитывая, что Т-супрессорные лимфоциты находятся в крови после трансплантации, а также их способность супрессировать экспрессию ко-стимулирующих молекул (CD80, CD86) на антигенпрезентирующих клетках донора, целесообразно проведение соответствующего мониторинга при трансплантации.
Как уже указывалось, изучение СD8+СD28-Т-лимфоцитов при аутоиммунной патологии также дает информацию о свойствах этих клеток. В частности, установлено, что у больных с красной системной волчанкой в активной фазе СD8+СD28-Т-лимфоциты не обладали ингибирующей активностью, что сочеталось с дисбалансом между ингибиторными влияниями IL-6 и стимулирующими IL-12.
По мнению авторов, нарушение баланса между таким ингибиторным цитокином, как IL-6 и стимулирующим IL-12 может быть причиной снижения функций супрессорных Т-лимфоцитов у этих больных.
Способность Т-супрессорных лимфоцитов подавлять пролиферацию антигенспецифических СD4+Т-лимфоцитов ассоциируется с появлением ремиссии у больных с аутоиммунной патологией. В системах in vitro удалось охарактеризовать предшественников CD8+CD28-T-лимфоцитов и показать, что ключевую роль в их генерации играют моноциты, которые секретируют IL-10 после стимуляции GM-CSF (в этих случаях прямой межклеточный контакт существенной роли не играет).
Предшественники имеют фенотип CD8+CD45RA-CD27-CCR-IL10Ra-. Было также показано, что Т-супрессорные лимфоциты подавляют активность и антигенспецифических цитотоксических лимфоцитов, уменьшая экспрессию антигенов I класса главного комплекса гистосовместимости.
Представленные данные не оставляют сомнений в том, что СD8+СD28-Т-лимфоциты играют важную роль в поддержании иммунологического гомеостаза вместе с СD4+-регуляторными Т-лимфоцитами.
Есть достаточно оснований считать, что одна из основных функций этих клеток — регуляция специфического Т-клеточного ответа. Активация супрессорных Т-лимфоцитов через специфическое распознавание в физиологических условиях защищает Th-лимфоциты от чрезмерной активации, а следовательно, и от чрезмерного иммунологического ответа в определенных условиях, в частности при увеличении количества реактивных Т-хелперов.
Имеющиеся данные показывают, что ингибирующие эффекты Т-супрессорных лимфоцитов являются одними из важных участников индукции периферической толерантности, а дисрегуляция контроля, который осуществляется этими клетками над аутореактивными клонами других Т-лимфоцитов, может быть причиной развития аутоиммунной патологии.
Наряду с этим нельзя не согласиться с тем, что понимание физиологического значения роли СD8+СD28-Т-лимфоцитов требует дальнейшего изучения, которое позволит получить новые данные о механизмах их действия в опухолевой и аутоиммунной патологии.
Общие представления о СD8+СD28-Т-лимфоцитах дает рис. 60.

Рис. 60. Фенотипические и функциональные особенности СD8+СD28-Т-лимфоцитов
Регуляторные функции супрессорных Т-лимфоцитов
Результаты изучения супрессорных СD8+СD28-Т-лимфоцитов не оставляют сомнений в том, что они выполняют важные регуляторные функции, которые достаточно четко определены в условиях нормы.
Вполне обоснованно также утверждать, что как супрессорные Т-лимфоциты (CD8+CD28-), так и регуляторные СD4+СD25+Т-лимфоциты (Th3/Trl) — совместные участники регуляции иммунологического гомеостаза на всех этапах его формирования.
Наряду с этим очевидно и то, что если роль супрессорных Т-лимфоцитов в условиях нормы достаточно ясна, то роль этих клеток при злокачественном росте предстоит выяснить в дальнейшем. В этом плане особый интерес представляет вопрос: при каких условиях CD8+CD28-T-cyпрессоры приобретают способность к цитотоксическому действию — факт, который наблюдался только при их культивировании с опухолевыми клетками.
Детализация этого вопроса (во всех ли случаях возможно появление способности к цитотоксичности, в какой мере это связано с биологическими особенностями опухолевых клеток, каков удельный вес этих клеток в реализации цитотоксичности и каков механизм влияния опухолевых клеток на СD8+СD28-Т-лимфоциты), несомненно, даст возможность нового понимания роли Т-супрессорных лимфоцитов в опухолевом процессе.
Не менее важно и дальнейшее изучение особенностей взаимодействия Т-супрессорных лимфоцитов с эндотелиальными клетками в процессе роста опухоли. Интерес к выяснению этого вопроса понятен в связи с тем, что взаимодействие СD8+СD28-Т-лимфоцитов с эндотелиальными клетками приводит к выраженным проявлениям активности последних, что может влиять на процесс злокачественной трансформации.
Обобщая представленные материалы, можно сделать такие выводы:
Первое
Т-супрессорные лимфоциты — CD8+CD28- представляют собой отдельный клон Т-лимфоцитов, экспрессирующих CD8, обладают выраженными ингибирующими эффектами в отношении СD4+Т-лимфоцитов, способны взаимодействовать с эндотелиальными клетками; выполняют важную роль в поддержании иммунологического гомеостаза в условиях нормы и патологии.
Второе
Ингибирующие эффекты супрессорных Т-лимфоцитов обусловлены межклеточными взаимодействиями, в которых наряду с СD8+СD28-Т-лимфоцитами принимают участие дендритные клетки и СD4+Т-лимфоциты.
Третье
В большинстве случаев при различных онкологических заболеваниях количество СD8+СD28-Т-лимфоцитов в крови увеличивается, что нередко сочетается с плохим прогнозом; лимфоциты, инфильтрирующие опухоль, также содержат значительное количество этих клеток.
Четвертое
При совместном культивировании Т-супрессорных лимфоцитов с аутологичными опухолевыми клетками появляются CD8+СD28-Т-лимфоциты, способные оказывать цитотоксическое действие.
Пятое
Определение количества СD8+СD28-Т-лимфоцитов может использоваться как контроль за влиянием химиотерапии.
Бережная Н.М., Чехун В.Ф.
Опубликовал Константин Моканов
Источник