Связь т и в лимфоцитов

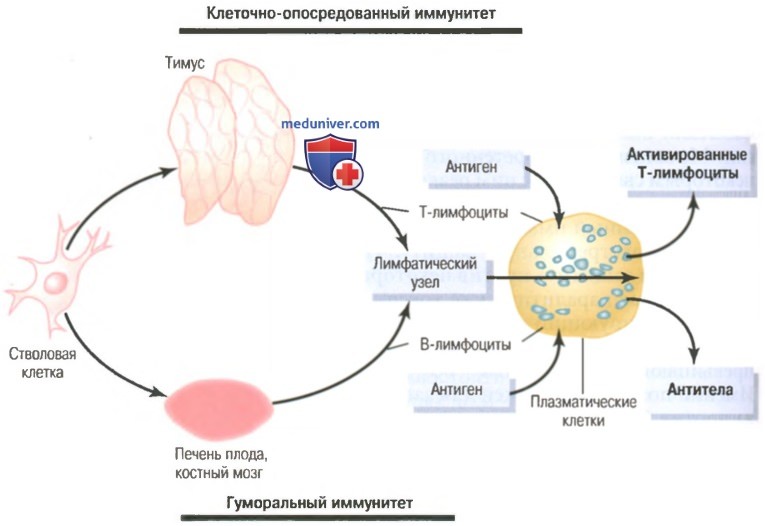
Т-лимфоциты и В-лимфоциты иммунитета. Предварительная обработка Т- и В-лимфоцитова) Клеточно-опосредованный и гуморальный иммунитеты обеспечивают два типа лимфоцитов – Т- и В-лимфоциты. Хотя при исследовании под микроскопом большинство лимфоцитов в нормальной лимфоидной ткани выглядят одинаковыми, эти клетки подразделяют на две основные популяции. Одна популяция — Т-лимфоциты — ответственна за формирование активированных лимфоцитов, обеспечивающих клеточно-опосредованный иммунитет. Другая популяция — В-лимфоциты — ответственна за формирование антител, обеспечивающих гуморальный иммунитет. Оба типа лимфоцитов образуются у эмбриона из полипотентных гемопоэтических стволовых клеток, формирующих лимфоциты как один из наиболее важных результатов их дифференцировки. Почти все сформированные лимфоциты в результате заселяют лимфоидную ткань, однако прежде чем это произойдет, они дополнительно дифференцируются или проходят предварительную обработку. Лимфоциты, которые в итоге станут активированными Т-лимфоцитами, сначала мигрируют к тимусу, где подвергаются предварительной обработке. Эти ответственные за клеточно-опо-средованный иммунитет лимфоциты называют Т-лимфоцитами, что подчеркивает роль тимуса. Другая популяция лимфоцитов, В-лимфоциты, предназначенные для формирования антител, проходят предварительную обработку в печени плода в середине периода внутриутробной жизни, а также в костном мозге в конце внутриутробной жизни плода и после рождения. Эта популяция клеток впервые была открыта у птиц, имеющих специальный орган для их предварительной обработки, который называют бурсой Фабриция (фабрициевой сумкой). Ответственные за гуморальный иммунитет лимфоциты называют В-лимфоцитами, что подчеркивает роль бурсы. На рисунке ниже показаны две лимфоцитарные системы для формирования: (1) активированных Т-лимфоцитов; (2) антител.
Предварительная обработка Т- и В-лимфоцитовВсе лимфоциты организма происходят от коммитированных в лимфоцитарном направлении стволовых клеток эмбриона, но эти клетки не могут непосредственно превратиться в активированные Т-лимфоциты или антитела. Прежде чем это станет возможным, клетки должны подвергнуться дальнейшей дифференцировке в соответствующих областях, где они проходят специфическую обработку. а) Т-лимфоциты проходят предварительную обработку в тимусе (вилочковой железе). После образования в костном мозге Т-лимфоциты сначала мигрируют к вилочковой железе. Здесь они быстро делятся, одновременно становясь чрезвычайно разнообразными, т.е. предназначенными для реакции против разных специфических антигенов. Это значит, что один лимфоцит, обработанный в тимусе, проявляет специфическую реактивность в отношении одного антигена. Следующий лимфоцит специфически реагирует на другой антиген. Это продолжается до тех пор, пока в тимусе не появятся тысячи разных типов лимфоцитов со специфической реактивностью в отношении тысяч разных антигенов. Эти разные типы предварительно обработанных Т-лимфоцитов оставляют тимус и распространяются кровью по всему телу, временно оседая в лимфоидной ткани. Кроме того, благодаря обработке в тимусе любой оставляющий его Т-лимфоцит не реагирует с белками или другими антигенами собственных тканей организма (иначе Т-лимфоциты погубили бы собственное тело человека в течение всего нескольких дней). Тимус выбирает, какие Т-лимфоциты могут его покинуть, сначала смешивая их практически со всеми специфическими аутоантигенами собственных тканей тела. Если Т-лимфоцит реагирует, он разрушается и фагоцитируется, вместо того, чтобы выделяться. Это происходит с основной частью клеток (вплоть до 90%). Таким образом, клетки, выделяющиеся из тимуса, не реагируют против собственных антигенов тела; они реагируют лишь на антигены внешних источников, например бактерий, токсинов или тканей, пересаженных от другого человека. Основная часть предобработки Т-лимфоцитов в тимусе происходит перед рождением ребенка и в течение нескольких месяцев после рождения. Удаление вилочковой железы после этого периода ослабляет (но не исключает) Т-лимфоцитарную иммунную систему. Однако удаление тимуса за несколько месяцев до рождения может нарушить развитие всего клеточно-опосредован-ного иммунитета. Поскольку именно клеточный тип иммунитета в основном отвечает за отторжение трансплантированных органов, например сердца или почек, органы можно пересаживать с меньшей вероятностью отторжения, если у животного в соответствующее время до его рождения удалить тимус. б) В-лимфоциты проходят предварительную обработку в печени и костном мозге. О деталях предварительной обработки В-лимфоцитов известно гораздо меньше, чем о предобработке Т-лимфоцитов. Известно, что у человека предварительная обработка В-лимфоцитов осуществляется в печени в середине внутриутробного периода развития, а также в костном мозге в конце внутриутробного периода и после рождения. Существуют два важных различия между В- и Т-лимфоцитами. Во-первых, В-лимфоциты активно секретируют реактивные агенты, называемые антителами, в отличие от Т-лимфоцитов, реагирующих с антигеном непосредственно. Антитела — это крупные белковые молекулы, способные соединяться с антигенной субстанцией и разрушать ее. Во-вторых, разнообразие В-лимфоцитов выражено больше, чем у Т-лимфоцитов, т.е. формируются миллионы типов В-лимфоцитарных антител с разными специфическими реактивностями. После предобработки В-лимфоциты, как и Т-лимфоциты, мигрируют к лимфоидной ткани по всему телу, где временно располагаются рядом, но несколько обособленно от областей локализации Т-лимфоцитов. – Также рекомендуем “Роль лимфоцитарных клонов. Происхождение клонов лимфоцитов” Оглавление темы “Клетки иммунитета. Виды иммунитета”: |
Источник
Взаимодействие Т- и В-лимфоцитов. Взаимодействие лимфоцитов и макрофаговНа основании всех изложенных данных были предложены различные гипотезы о трехклеточных системах иммунопоэза. В 1969 г. несколько авторов независимо друг от друга предположили, что процесс антителогенеза инициируется в результате кооперации трех типов клеток: клеток-предшественников (В-лимфоцитов), Т-лимфоцитов и макрофагов. При возникновении антигенного стимула В-лимфоцит-предшественник под влиянием сигналов со стороны Т-клеток-помощников и макрофагов переходит из покоящегося состояния в метаболически активную фазу, обеспечивающую клональную пролиферацию и дифференцировку в плазматическую клетку, активно синтезирующую и секретирующую антитела. Роль макрофагов существенна, по-видимому, на самом первом этапе распознавания антигена и приведения его в иммуногенную форму. Опыты с использованием антимакрофагальных сывороток показали необходимость присутствия макрофагов лишь в первые сутки после введения антигена, т. е. в момент антигенной стимуляции (Галактионов, Анфалова, 1974). Все теории индукции антителогенеза в В-лимфоцитах можно разделить на две группы: теория односигнальной и двухсигнальной активации В-лимфоцнтов. Сторонники односигнальных теорий постулируют необходимость одного сигнала для перехода покоящегося В-лимфоцита в метаболически активную фазу. Одни авторы считают, что этим сигналом является неспецифическое ми-тогенное влияние антигена, а не взаимодействие антигенных детерминантов с рецепторами на поверхности В-клетки. Иммуноглобулиновые рецепторы, по их мнению, обеспечивают лишь концентрирование антигена на определенных В-клетках, что приводит к преимущественной активации клонов, детерминированных к синтезу антител данной специфичности.
Экспериментальное обоснование этой гипотезы опирается на результаты опытов но изучению влияния тимуснезависимых антигенов на В-клетки. Большинство известных тимуснезависимых антигенов обладают свойствами поликлональных активаторов и являются митогенами для В-клеток (Doenhoff е. а., 1976). Имеются данные о том, что некоторые В-клеточные митогены вызывают поликлона льный синтез антител в отсутствие специфического антигена (Andersson е. а., 1972). Отсюда делается вывод, что сигналом, вызывающим индукцию иммунной реактивности в В-клетке, является неспецифическое митогенное воздействие, а не связывание специфических антигенных детерминант иммуноглобулиновыми рецепторами. Однако есть факты, которые не укладываются в представление о ведущей роли поликлонального воздействия в активации В-клетки. Так, существуют Т-независимые антигены, которые не вызывают митогенного эффекта или поликлонального синтеза антител, Кроме того, есть сведения об отсутствии корреляции между степенью митогенности тимуснезависимого носителя н силой иммунного ответа к присоединенному к нему гаптену. Известно, например, что пневмококковый полисахарид является лучшим митогеном, чем леван, однако ответ к динитрофенильной группе, конъюгированной с леваном, был выше, чем при конъюгировании данного гаптена с пневмококковым полисахаридом. Вторая точка зрения о природе стимулирующего сигнала заключается в том, что активация В-клеток происходит в результате взаимодействия антигенных детерминант с иммуноглооулиновыми рецепторами В-лимфоцита (Feldmann e. a., 1975 b; Klaus, Hamphrey, 1975). В случае тимуснезависимых антигенов многоточечное присоединение густо повторяющихся детерминант к поверхности В-клетки обеспечивает высокую энергию связывания антигена с В-лимфоцитом, что приводит к его активации. В случае тимуезависимых антигенов необходимая для активации структура антигенных детерминант создается с помощью Т-лимфоцитов, которые концентрируют антиген и представляют его В-клетке в поливалентной форме. Так, согласно гипотезе Фелдмана и соавторов, Т-клетки полимеризуют антиген на поверхности макрофага. Молекулы антигена связываются носителем с IgT, вырабатываемым активированными Т-лимфоцитами. Комплексы антиген — IgT присоединяются к макрофагу, в результате чего на поверхности макрофага создается обойма из антигенных молекул, ориентированных гаптенными группировками наружу. Такая структура с высокой плотностью идентичных антигенных детерминант и активирует В-клетку. При отсутствии макрофагов, являющихся местом структурной организации антигена, развивается не иммунный ответ, а толерантность. Таким образом, односигнальная модель также предусматривает кооперацию различных типов клеток при создании активирующего стимула. Существование тимуснезависимых антигенов, очевидно, не противоречит положению о необходимости взаимодействия клеток при индукции антителогенеза. – Также рекомендуем “Модель активации В-клетки. Индукция антителообразования” Оглавление темы “Супрессия иммунного ответа. Клетки супрессоры”: |
Источник
Варианты взаимодействия $T$- и $B$-лимфоцитов при иммунном ответе
Единственными эффективными антигенпрезентирующими клетками при первичном иммунном ответе для $T$-лимфоцитов являются дендритные клетки. Если при активации $T$-лимфоцита антиген был презентирован дендритными клетками, то в иммунный ответ вовлекаются $B$-лимфоциты, расположенные рядом.
Возможны следующие варианты взаимодействия $T$- и $B$-лимфоцитов:
- $B$-лимфоциты связывают растворимый антиген через BCR, путем эндоцитоза его поглощают, подвергают процессингу и на поверхности экспонируют фрагменты антигена совместно с молекулами МНС-II. Антиген связывается на поверхности $B$-лимфоцита TCR в комплексе с МНС-II. Устанавливаются необходимые корецепторные связи между $T$- и $B$-лимфоцитами. Такое взаимодействие наблюдается в $T$-зависимых зонах периферической лимфоидной ткани в самом начале развития иммунного ответа.
- Специфический антиген распознается $B$-лимфоцитом и вблизи находится активированный $T$-лимфоцит, который распознал антиген на другой антигенпрезентирующей клетке. Взаимодействие $T$- и $B$-лимфоцитов может проявиться восприятием цитокиновых сигналов $B$-клеткой от $B$-лимфоцита.
Замечание 1
Взаимодействие с участием мембранных молекул при первичном иммунном ответе может наступать или нет. При вторичном иммунном ответе принимают участие $T$-клетки памяти. В обязательном порядке происходит взаимодействие мембранной молекулы $B$-лимфоцита $CD40 с$ мембранной молекулой $T$-клетки $CD40L$. Вторичный ответ $B2$-лимфоцитов происходит с обязательным переключением изотипа иммуноглобулинов с $IgM$ на $Iga$, $IgG$, $IgE$. Эти взаимодействия происходят в фолликулах лимфоидных органов (в $B$-клеточных зонах).
Реализация реакции иммунной системы на антиген
После внедрения антигена во внутреннюю среду организма наблюдается его фагоцитирование или поглощение макрофагами. Макрофаги с помощью протеолитических ферментов убирают детерминантные группировки чужих макромолекул. Образовавшиеся детерминанты в результате экзоцитоза попадают на поверхность мембраны, где соединяются с мембранными белками макрофага.
Белки макрофага кодируются генами главного комплекса гистосовместимости, которые обеспечивают синтез белков, определяющих иммунологическую индивидуальность организма.
Антигены гистосовместимости (мембранные белки) являются трансплантационными антигенами, так как различны у разных индивидов. Эти антигены вызывают реакцию отторжения трансплантата при пересадке тканей и органов.
Антигены гистосовместимости представляют $T$-лимфоцитам очищенную антигенную детерминанту.
Передача антигена с макрофага $T$-лимфоциту происходит в результате следующих процессов:
- $T$-лимфоцит определяет чежеродность детерминантной группы антигена, находящегося на поверхности макрофага;
- рецептор $T$-лимфоцита связывается с антигеном;
- происходит активация $T$-лимфоцита;
- в $T$-лимфоците образуется белок – интерлейкин-2;
- на мембранах $T$-клеток формируются рецепторы к интерлейкину, происходит связывание с ним;
- начинаются процессы деления и дифференцировки $T$-клеток с образование клеток-эффекторов (посредники при вовлечении в иммунный ответ $B$-лимфоцитов, $T$-хелперы) или клонов ($T$-киллеры).
$T$-лимфоциты-хелперы при участии макрофагов передают информацию об антигене $B$-клеткам. Роль макрофагов в данном процессе:
- снимают антигенрецепторные комплексы с поверхности $T$-лимфоцитов;
- концентрируют на своей поверхности эти комплексы в виде «обоймы»;
- передают $B$-лимфоциту «обоймы» антигенных молекул.
Для активации $B$-лимфоцита необходимо, чтобы молекула антигена была подготовлена макрофагом и $T$-лимфоцитом. Таким образом происходит передача специфической информации об антигене от $T$-лимфоцита $B$-лимфоциту.
В виде гуморального стимула передается второй неспецифический сигнал от $T$-клеток $B$-лимфоцитам.
Замечание 2
$B$-лимфоциты, после получения специфического и неспецифического сигналов, пролиферируют с дифференцировкой в различные клоны эффекторных клеток. При этом образуются плазматические клетки, продуцирующие иммуноглобулины.
Источник
Оглавление темы “Селезенка. Лимфатические узлы. Стадии иммунного ответа. Формы иммунного ответа. Воспаление. Механизмы, контролирующие иммунную систему.”: Активация Т- и В-лимфоцитов в иммунном ответе. Активация лимфоцитов. Формы специфического иммунного ответа.Передача сигнала активации от антигенраспознающего рецептора в ядро клетки приводит к активации конкретных генов, ответственных за митотическую активность клеток, контролирующих продукцию клеткой биологически активных молекул, участвующих в антимикробной защите (антитела, цитокины).
В-лимфоцит получает одновременно два сигнала активации: от антигенраспознающего рецептора при его соединении с антигеном и от связывания его поверхностных костимулирующих молекул с соответствующими лигандами на Т-лимфоцитах. После этого В-лимфоцит пролиферирует и потомки его превращаются в зрелые антителопродуцирующие плазматические клетки. Т-лимфоцит в ответ на контакт с антигеном начинает пролиферировать, потомки его приобретают способность продуцировать определенные цитокины или превращаются в зрелые цитотоксические клетки. В зависимости от того, какие дополнительные сигналы активации (цитокины, костимулирующие молекулы) получает Т-лимфоцит в момент контакта с антигеном, его потомки дифференцируются в двух разных направлениях, превращаясь в Т-хелперы, продуцирующие гамма-интерферон (Th1), или в Т-хелперы, продуцирующие интерлейкины-4, -5, -6, -10, -13 (Th2). Количественное преобладание Thl над Th2 является условием развития клеточного (кле-точно-опосредованного) иммунного ответа. В случае преобладания Th2 развивается гуморальный иммунный ответ, проявляющийся продукцией специфических антител (рис. 8.9). – Также рекомендуем “Клеточный иммунный ответ. Гуморальный иммунный ответ. Защитные функции иммуноглобулинов ( антител ).” |
Источник