Связь с кровью понятие о рециркуляции лимфоцитов

Лимфоциты постоянно рециркулируют из крови в ткани, и обратно. Рециркуляцией называют процесс перехода лимфоцитов из кровяного русла в органы, оттуда в лимфу и вновь в кровоток. При этом клетки, покинувшие конкретный лимфатический узел, возвращаются в любой узел или селезенку, что и обеспечивает перемешивание рециркулирующих лимфоцитов.
Лимфоциты переходят из кровотока в лимфоидные образования обычные посткапиллярные венулы, однако, этот переход обычно осуществляется преимущественно через специализированные участки венозного русла – венулы с высоким эндотелием. В лимфоузлах эти сосуды находятся главным образом в паракортикальной области и иногда в корковой. Вместе с тем часть лимфоцитов, а в первую очередь Т-клетки, поступают в региональный лимфатический узел из дренируемой им области по приносящим лимфатическим сосудам.
Из лимфатических узлов лимфоциты возвращаются в кровоток по приносящим лимфатическим сосудам через грудной поток в краниальную полую вену. Ежечасно в рециркуляцию вовлекается 1-2% лимфоцитов, большинство зрелых лимфоцитов поступает из органов в циркуляцию и обратно, причем, время, соответствующее половине срока их однократного пребывания в циркуляции, составляет до 30 минут. В итоге этот процесс позволяет множеству антигенспецифичных лимфоцитов встретиться с соответствующими антигенами, проникшими в периферические лимфоидные органы. Особая важность рециркуляции становится очевидной, если учесть, что лимфоциты моноспецифичны и лишь ограниченное число лимфоцитов способно распознавать каждый конкретный антиген.
В норме рециркуляция лимфоцитов через лимфатические узлы происходит постоянно. Однако, если в лимфатический узел, ранее сенсибилизированный к конкретному антигену, повторно попадает антиген, то рециркуляция прекращается примерно на 24 часа. Временная остановка рециркуляции обусловлена избирательной задержкой антигенспецифичных лимфоцитов в лимфоузлах, дренирующих место проникновения антигена. Лимфоциты уже не рециркулируют, а остаются в участке встречи с антигеном.
Специфическая рециркуляция лимфоцитов происходит в лимфоидной ткани, ассоциированной со слизистыми оболочками. Лимфоциты этой ткани возвращаются в процессе рециркуляции образом в эту же ткань. Так, лимфоциты, стимулированные в пейеровых бляшках, проходят через региональные лимфатические узлы в кровоток, а затем возвращаются «домой» в собственную пластинку слизистой оболочки. Такая специфическая рециркуляция объясняется тем, что эти лимфоциты экспрессируют молекулы «возврата домой», которые связываются со специальными молекулами адгезии-адрессиинами на поверхности эндотелиальных клеток венул лимфоидной ткани слизистых оболочек, что и обеспечивает избирательную рециркуляцию. По этой же причине стимуляция антигеном в области слизистой оболочки в том или ином участке организма вызывает образование антител преимущественно в слизистых оболочках.
В определении направления миграции лимфоцитов и в преодолении барьеров между кровью и тканями основную роль играют хемотаксические сигналы и специфические межклеточные взаимодействия, основанные на взаимном распознавании мембранных структур клеток.
Источник
Лимфоциты
периферической крови мигрируют через
посткапиллярные венулы в лимфатические
узлы или селезенку. Т-клетки заполняют
перифолликулярные зоны кортикальных
областей лимфатических узлов
(паракортикальные области) и
периартериолярные тяжи, окружающие
центральные артериолы селезенки.
В-клетки селективно аккумулируются в
зародышевых фолликулах лимфатических
узлов и селезенки, а также в субкапсулярной
зоне коры и медуллярных тяжах лимфатических
узлов. Лимфоциты возвращаются в
периферическую кровь эфферентным током
лимфы через грудной лимфатический
проток. Большинство рециркулирующих
клеток (в грудном лимфатическом протоке
или периферической крови) являются
Т-клетками, а средняя продолжительность
их полной циркуляции составляет около
10 часов. В норме в периферической крови
и зародышевых центрах преобладают
хелперные клетки CD4+,
а в костном мозге и лимфоидной ткани,
ассоциированной с кишечником, преобладают
Т-лимфоциты CD8+.
Т-клетки, экспрессирующих gd-рецептор
мигрируют преимущественно в лимфоидную
ткань, ассоциированную с легкими, кожей
и кишечником. Большинство В-клеток ведет
более “оседлый” образ жизни, проводя
длительное время в селезенке и
лимфатических узлах.
Список литературы
Кетлинский
С.А., Калинина Н.М. Иммунология для врача.
– С-Пб.: Гиппократ, 1998. – 156с.Кетлинский
С.А., Симбирцев А.С., Воробьев А.А.
Эндогенные иммуномодуляторы.- СПб:
Гиппократ, 1992. – 256 с.Фрейдлин
И.С. Иммунная система и ее дефекты.
Руководство для врачей. СПб.:НТФФ
Полисан, 1998. – 112с.Ярилин
А.А. Система цитокинов и принципы ее
функционирования в норме и при патологии
// Иммунология. – 1997. – №5. – С.7-14Bona
C., Bonilla F. Textbook of immunology, second ed., Harwood Acad.
Publ., Amsterdam, 1996, 406p.Boyd
J., Tucek C., Godfrey D. Et al. The thymic microenvironment //
Immunol. Today. – 1994. – Vol. 14. – P.445 – 449Dorshkind
K.
Lymphopoiesis
/
InHoffman
R., Benz E.J., Sanford J. et al. Hematology Basic Principles and
Practice: Churchill Livingstone, London. -1995. -2325 р.Mac
Lennan I.C.M. Germinal centers // Ann. Rev. Immunol. –1994. –
Vol.12. – P.117 – 139Schwartz
J. A. and Schwartz R. S. Structure and Function of the Immune System
/ InHoffman
R., Benz E.J., Sanford J. et al. Hematology Basic Principles and
Practice: Churchill Livingstone, London.
-1995. -2325 р.
Таблица
4.2.1.
Антигены
дифференцировочных кластеров (CD)
лейкоцитов
Антигены | Функциональные | Экспрессия |
CD2 | Рецептор | Т-клетки, |
CD3 | Комплекс | Все |
CD4 | Лиганд | Т-хелперы |
CD5 | Лиганд | Т-клетки, |
CD7 | Рецептор | Ранние |
CD8 | Лиганд | Цитотоксические |
CD9 | Молекула | Пре-В-клетки, |
CD10 | Нейтральная | Про-В-клетки, |
CD11a CD11b CD11c | ?–цепь ?–цепь Рецептор | Лейкоциты Гранулоциты, Моноциты, |
CD13 | Аминопептидаза | Гранулоциты, |
CD14 | Мембранный | Моноциты, |
CD16 | Рецептор | NK-клетки, |
СD19 | В-клеточный | Ранние |
СD20 | ? | Ранние |
Продолжение | ||
Антигены | Функциональные | Экспрессия |
CD21 | Рецептор | В-клетки, |
CD22 | Принадлежит | В-лимфоциты |
CD23 | Fc? | Субпопуляция |
CD25 | Низкоаффинный | Активированные |
CD28 | Лиганд | Субпопуляция |
СD30 | Ki-1, | Т-клетки, |
CD32 | Рецептор | Субпопуляция |
CD33 | ? | Миелоидные |
CD34 | Лиганд | Плюрипотентная |
CD40 | Лиганд | В-клетки, |
СD44 | Лиганд | Рецептор, |
CD45RO CD45RA,В | Лиганд | Активированные “наивные” |
CD56 | Адгезионная | NK-клетки |
Таблица
4.2.2.
Этапы
дифференцировки В-лимфоцитов
Определяющие | Костный | Лимфоидная | |||
Про-В-клетка | Большая | Малая | Зрелая (наивная) | Активированная | |
Дифференцировочные | HLA-DR CD34 CD19 CD10 CD40 CD22 | HLA-DR CD19 CD10 CD40 CD22 | HLA-DR CD19 CD22 CD20 CD21 (CD5)* | HLA-DR CD19 CD20 CD22 CD21 | HLA-DR CD19 CD20 CD40 CD22 |
Перестройка | Не | Реаранжировка | Реаранжировка | Антигензависимая | Антигензависимая |
Иммуноглобулиновый | Пре-R | Пре-R | Пре-R | sIgM, sIgD | sIgM |
Регулирующие | Строма | Аутоантигены | Чужеродные | ||
Примечание: Про
– наиболее ранний предшественник
В-клеток; Пре – предшественник В-клетки
ПреR
– предшественник В-клеточного рецептора;
sIgM,
sIgD – поверхностные формы иммуноглобулинов
М и D; * – маркер субпопуляции В1а
Таблица
4.2.3.
Этапы
дифференцировки Т-лимфоцитов
Определяющие | Про-Т- ТН | Пре-T-клетка ТН | Тимоцит ТН | Тимоцит ДП | Зрелые |
Дифференцировочные | CD3- СD4- CD8- | CD3?? СD4- CD8- | CD3??комплекс СD4- CD8- | CD3 СD4+,CD8+ | CD3 СD4 CD8 |
.Перестройка | Реаранжировка | Реаранжировка | TCRa?? Или TCR?? | ||
Расположение | Тимус, | Тимус, | Тимус, | Тимус, | Лимфоидные |
Важнейшие Регулирующие | Активная | Активная | Контакт | Контакт | Цитокины, |
Примечание:
ТН – трижды негативные (CD3-,4-,8-);
ДП – дубльнегативные(CD4-,8-)


Рис.4.2.3.
Экспрессия некоторых поверхностных
молекул при развитии Т-клеток
Рис.4.2.4.
Пути созревания В-лимфоцитов в
лимфатическом узле после выхода из
костного мозга

Соседние файлы в папке Внутренние
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Оглавление темы “Иммунная система. Функции клеток иммунной системы. Т-лимфоциты. В-лимфоциты. Структура и функции органов иммунной системы.”: Иммунная система. Иммунная система человека. Неспецифический иммунитет. Специфический иммунный ответ. Рециркуляция лимфоцитов.Во внутренней среде организма присутствуют клетки и молекулы, которые обеспечивают защитную функцию (иммунитет). Часть из них относятся к факторам врожденного иммунитета, т. е. присутствуют в организме еще до встречи с каким-либо болезнетворным микроорганизмом или чужеродной молекулой. К таким факторам защиты относятся фагоцитирующие клетки крови и тканей, а также большие гранулярные лимфоциты, получившие название «натуральные (естественные) киллеры». Врожденными факторами защиты организма являются также молекулы, продуцируемые и секретируемые вышеназванными клетками, а также лимфоцитами, стромальными и другими клетками. К таким молекулам относятся: белки системы комплемента, медиаторы межклеточного взаимодействия — цитокины и др. Белки системы комплемента продуцируются постоянно и циркулируют в крови, большинство цитокинов начинают продуцироваться и секре-тироваться клетками только в ответ на сигнал активации (например, на контакт с микроорганизмом). Более эффективным способом защиты внутренней среды организма от проникающих в нее чужеродных агентов (антигенов) является специфический иммунный ответ, в результате которого организм приобретает дополнительные защитные механизмы: активированные клетки и продуцируемые ими молекулы. Защитное действие этих механизмов строго избирательно (специфично) в отношении того конкретного антигена (например, патогенного микроорганизма), контакт с которым вызвал иммунный ответ. Специфический иммунный ответ является функцией клеток и органов иммунной системы.
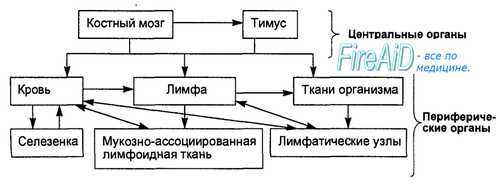
Главными клетками иммунной системы являются Т- и В-лимфоциты, которые рециркулируют в системе кровотока и лимфотока, постоянно перемещаясь из одних органов иммунной системы в другие, обладают способностью выходить в ткани для выполнения защитных функций (рис. 8.1). В защитных реакциях специфического иммунитета кроме Т- и В-лим-фоцитов участвуют фагоцитирующие клетки (гранулоциты, моноциты, макрофаги), «естественные киллеры, тучные клетки, эндотелиальные и эпителиальные клетки, которые играют роль вспомогательных, взаимодействуя с Т- и В-лимфоцитами. – Также рекомендуем “Происхождение ( образование ) клеток иммунной системы. Функции клеток иммунной системы. Лимфопоэз. Бурса Фабрициуса..” |
Источник
Лимфоидные органы — динамичные образования: лимфоциты непрерывно перемещаются в стромальном каркасе, покидают лимфоидные органы и вновь возвращаются в них. Этот круговорот лимфоцитов обозначают термином «рециркуляция». В процессе рециркуляции лимфоциты покидают лимфоидные органы с эфферентной лимфой (из селезенки они выходят с кровью). Через грудной лимфатический проток лимфоциты проникают в кровяное русло. Затем через посткапиллярные венулы клетки вновь проникают в лимфоидный орган (тот же или другой), внутри которого перемещаются в специфические места их локализации. Затем клетки вновь покидают орган и выходят в циркуляторное русло и т.д. В нелимфоидных органах лимфоциты практически не циркулируют.
Из сказанного следует, что выявляемые в крови лимфоциты находятся в ней недолго (примерно 30 мин), но в течение суток многократно (4—5 раз) покидают ее и вновь возвращаются. Таким образом, несмотря на то, что в крови лимфоциты составляют всего около 0,5% от общего числа клеток, их состояние достаточно точно отражает состояние популяции лимфоцитов в целом организме. Это очень существенно, поскольку именно лимфоциты крови человека служат наиболее обычным и доступным объектом изучения как в экспериментальных, так и в клинико-иммунологических исследованиях. Субпопуляционный состав лимфоцитов крови человека представлен в табл. 3.23.
Таблица 3.23. Нормальные показатели содержания в крови взрослых людей основных популяций и субпопуляций лимфоцитов (по С.В. Хайдукову, 2008)
| Клетки | Относительное содержание*, % | Число клеток в 1 л (М+m) |
| Лимфоциты (CD45+)** | 32+4 | 1,363—2,808х109 |
| В-клетки ^D3-CD19+) | 12+5 | 0,111—0,376х109 |
| Т-клетки (CD3+CD19-) | 73+12 | 0,946-2,079х109 |
| арТ-клетки (CD3+apTCR+) | 70,5+9,7 | 0,022—0,115х109 |
| yST-клетки (CD3+ySTCR+) | 4,6+2,8 | 0,924—1,964х109 |
| Т-хелперы (CD3+CD4+) | 45+10 | 0,576—1,336х109 |
| Т-киллеры (CD3+CD8+) | 27+8 | 0,372—0,974х109 |
| Регулятоные T-клетки (CD3+CD4+CD25hi) | 3,7+2,05 | 0,009—0,078х 109 |
| NKT-клетки (CD3+CD56+CD16+) | 3+3 | 0,007—0,165х109 |
| Т-хелперы наивные (CD3+CD4+CD45RA+) | 30+10 | 0,272—1,123х109 |
| Т-хелперы активированные/памяти (CD3+CD4+CD45R0+CD29+) | 15+10 | 0,068—0,702х109 |
| NK-клетки (CD45+CD56+СD16+) | 13+5 | 0,123—0,369х109 |
* Указано среднее арифметическое со стандартной ошибкой.
** Относительные и абсолютные количества лимфоцитов определяли от общего количества лейкоцитов.
Относительные и абсолютные количества субпопуляций Т-, В- и NK-клеток определяли от общего количества лимфоцитов.
Способность клеток находить «свое место» в организме называют хомингом. Существует две группы механизмов, задействованных в данном процессе, — контактные механизмы, обеспечиваемые молекулами адгезии, находящимися на поверхности лимфоцитов, и хемотаксис, определяющий направленность движения клеток, зависящую от наличия на их поверхности хемокиновых рецепторов.
Попав в сосудистую сеть лимфоидного органа, лимфоциты реагируют на хемокины, вырабатываемые клетками эндотелия в специализированном для миграции лимфоцитов отделе сосудов — посткапиллярных венулах.
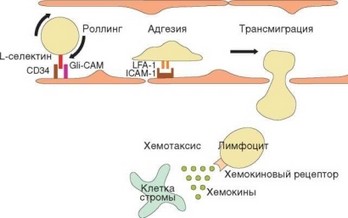
Рис. 3.74. Схема транссосудистой миграции лимфоцитов. Отражены основные стадии эмиграции лимфоцитов через высокий эндотелий посткапиллярных венул лимфоидных органов. Указаны молекулы адгезии, важные для осуществления этого процесса; отражена роль хемокинов в привлечении клеток
Только в посткапиллярных венулах лимфоидных (но не других) органов есть высокий эндотелий, образованный спонтанно активированными клетками (аналогичный высокий эндотелий может быть индуцирован в других органах только под влиянием провоспалительных факторов). Клетки этого эндотелия секретируют р-хемокин ССЫ9, распознаваемый рецептором CCR7. Этот рецептор экспрессируют на поверхности наивные лимфоциты как Т-, так и В-класса. Таким образом, указанные эндотелиальные клетки способны привлекать как Т-, так и В-лимфоциты.
Далее происходит трансмиграция лимфоцитов по механизму, аналогичному миграции лейкоцитов в очаг воспаления (отличия касаются конкретных молекул адгезии, определяющих направление миграции — рис. 3.74). Ключевую роль при этом играет начальный момент миграции, обусловленный взаимодействием мембранного L-cелектина (СD62L) лимфоцитов с адрессинами (GlyCAM, CD34, PSGL-1) эндотелиальных клеток. Именно наличие CD62L на клеточной мембране определяет способность наивных лимфоцитов мигрировать в лимфатические узлы и пейеровы бляшки, в связи с чем CD62L называют рецептором хоминга. Это слабое взаимодействие обусловливает феномен качения (перекатывания лимфоцита вдоль сосудистой стенки). Вслед за этим устанавливается более прочное взаимодействие между р2-интегрином лимфоцитов LFA-1 и его рецептором на эндотелиальной клетке — ICAM-1. Взаимодействие усиливается благодаря «активации» р2-интегрина (т.е. изменения конформации молекулы, в результате которого повышается его сродство к рецептору — см. раздел 2.3.1.2). Это обусловливает прочное прилипание лимфоцита к сосудистой стенке. Затем под влиянием хемотаксических сигналов, поступающих из ткани
лимфоидного органа, лимфоцит начинает мигрировать из сосуда между эндотелиальными клетками.
Вышедшие из сосуда клетки попадают в примыкающие к посткапиллярным венулам тимусзависимые зоны, стромальные и дендритные клетки которых секретируют хемокины CCL19 (ELC) и С^21 (SLC), распознаваемые рецептором CCR7 на лимфоцитах. До этого момента Т- и В-клетки перемещаются в одном и том же направлении. После того как лимфоцит попадает во внутреннюю среду органа, их пути расходятся. Т-клетки остаются в тимусзависимой зоне (в паракортексе лимфатических узлов, параартериальной муфте белой пульпы селезенки и Т-зонах пейеровых бляшек). В-клетки реагируют на хемокин CXCR13 (BLC), к которому они экспрессирут рецептор СХСR5. Этот хемокин секретируют стромальные клетки фолликулов, что и определяет направление миграции В-лимфоцитов. В-лимфоциты, не несущие СХСR5, не поступают в фолликулы и локализуются в прилежащей к фолликулу зоне. Механизмы, определяющие дальнейшее перемещение лимфоцитов из органа в эфферентную лимфу, пока не установлены.
В процессе рециркуляции отсутствует «привязка» хоминга к конкретному лимфатическому узлу или пейеровой бляшке. Так, лимфоцит, покинувший шейный лимфоузел, может затем проникнуть, например, в паховый лимфатический узел или в пейерову бляшку. Принципиально лишь то, что клетки не выходят из системы вторичных лимфоидных органов. Степень специфичности хоминга минимальна в селезенке, так как лимфоциты проникают в белую пульпу при излиянии крови в краевой зоне. Только последующее перемещение в Т- и В-зоны осуществляется по описанному выше механизму.
Все сказанное выше относится к наивным лимфоцитам. После активации лимфоцитов антигеном экспрессия молекул адгезии и хемокиновых рецепторов на их поверхности существенно изменяется. Это определяет иные пути миграции клеток после контакта с антигеном. Активированные лимфоциты и клетки памяти очень слабо экспрессируют L-селектин, что обусловливает низкую вероятность их попадания в лимфатические узлы. В то же время они приобретают способность мигрировать в очаги воспаления, расположенные в нелимфоидных органах, а также в барьерные ткани (рис. 3.75). Пути миграции активированных лимфоцитов и клеток

Рис. 3.75. Мембранный фенотип Т-клеток определяет пути их миграции. Указаны молекулы адгезии, определяющие различные направления миграции Т-клеток
памяти будут специально рассмотрены далее (см. раздел 3.6.5.4). Благодаря различной экспрессии рецепторов хоминга в разных тканях формируется несколько относительно автономных путей рециркуляции. Пути рециркуляции наивных лимфоцитов и клеток памяти могут пересекаться. Так, и наивные клетки, и клетки памяти, активированные в кишечнике, попадают в брыжеечные лимфатические узлы.
Интенсивность рециркуляции Т-лимфоцтов существенно выше, чем В-лимфоцитов. У человека через каждый лимфатический узел за сутки проходит 0,3х10п, через селезенку — 2,5х10п, а через кровь — 5х10п лимфоцитов. Длительность пребывания Т-клеток в кровотоке за 1 цикл рециркуляции — около 30 мин. Благодаря рециркуляции происходит постоянное «перемешивание» лимфоидных клеток с сохранением описанного выше порядка: наивные клетки, как правило, не покидают лимфоидной ткани, тогда как клетки памяти, наоборот, часто выходят за ее пределы, мигрируя, например, в эпителиальные органы и ткани, а также в очаги воспаления. Внутри лимфоидных органов клетки также занимают соответствующие места. Упорядоченность локализации лимфоцитов различных типов очень важна для развития иммунного ответа.
Источник