Ретикулярная ткань стромы лимфатического узла

Лимфатические узлы. Развитие лимфатических узлов. Строение лимфатических узлов.
Лимфатические узлы — это органы, расположенные по ходу лимфатических сосудов. Размер их 0,5-1 см, форма — чаще округлая, овальная или бобовидная. Располагаются они обычно регионарно, группами. С выпуклой стороны узла в него входят приносящие лимфатические сосуды, а с противоположной, называемой воротами, выходят выносящие лимфатические сосуды. Кроме того, в ворота входят артерии и нервы и выходят вены. Общее число лимфатических узлов достигает 1 тыс., что составляет около 1% массы тела.
Лимфатические узлы выполняют роль активного биологического фильтра, в котором задерживается и фагоцитируется до 99% всех инородных частиц и бактерий.
Различают неспецифическую защитную функцию лимфатических узлов за счет элиминации микробов из лимфы и специфическую, выражающуюся в иммунном ответе на антигены. Эти органы выполняют и кроветворную функцию. Хотя стволовые клетки в них практически отсутствуют, но пролиферация лимфобластов, дифференцировка В-лимфоцитов в плазмоциты происходит. Лимфа, протекая через лимфатические узлы, обогащается лимфоцитами.
Развитие лимфатических узлов. Зачатки лимфатических узлов появляются в конце 2-го – начале 3-го месяца эмбриогенеза в виде скоплений мезенхимы по ходу лимфатических сосудов. Вскоре из мезенхимы образуется ретикулярная ткань, составляющая строму органа. К концу 4-го месяца в закладки узлов вселяются лимфоциты и формируются скопления — первичные узелки без центра размножения. Одновременно появляется подразделение органа на корковое и мозговое вещество.
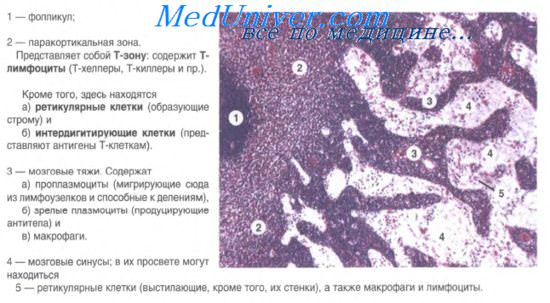
Строение лимфатических узлов. В лимфатическом узле имеются следующие структурные компоненты: капсула, содержащая много коллагеновых волокон (в области ворот в капсуле есть и гладкие миоциты), трабекулы — перекладины из соединительной ткани, которые, анастомозируя друг с другом, образуют каркас узла, ретикулярная ткань, заполняющая все пространство, ограниченное капсулой и трабекулами.
В лимфатическом узле различают периферическое корковое вещество и центральное мозговое вещество. Между этими частями выделяют еще паракортикальную зону.
В корковом веществе расположены скопления лимфоидной ткани в виде вторичных узелков. Это округлые образования диаметром до 1 мм. Центральную часть узелка называют центром размножения, или реактивным центром. Здесь происходит антигензависимая пролиферация В-лимфоцитов и дифференцировка их в предшественники плазматических клеток. Кроме того, в центре размножения находятся дендритные клетки костномозгового происхождения, которые на своих отростках удерживают антигены, активирующие В-лимфоциты, макрофаги моноцитарного генеза, фагоцитирующие погибающие апоптозом аутоиммунные В-лимфоциты, антигены и инородные частицы.
По периферии вторичного узелка расположена корона полулунной формы, состоящая из малых лимфоцитов (рециркулирующих В-лимфоцитов, В-клеток памяти, незрелых плазматических клеток). На границе центра размножения короны обнаруживаются Т-лимфоциты (хелперы), которые способствуют развитию В-лимфоцитов в иммунобласты. Последние мигрируют в мозговые тяжи, отходящие от паракортикальной зоны и узелков внутрь мозгового вещества.
Лимфатические узелки являются динамическими структурами. Они то образуются, то исчезают. В процессе изменения структуры лимфатических узелков различают несколько стадий (формирование светлого центра размножения, появление вокруг центра короны из малых лимфоцитов и др.). Период их существования — 2-3 су т.
Паракортикалъная зона лимфатического узла находится на границе между корковым и мозговым веществом. Она называется тимусзависимой зоной, или Т-зоной, так как при удалении тимуса происходит ее исчезновение. В паракортикальной зоне осуществляются бласттрансформация Т-лимфоцитов, их пролиферация и превращение в специализированные клетки системы иммунитета. Здесь много дендритных клеток. Они появляются в результате миграции из тканей системы покрова организма внутриэпидермальных макрофагов. На своей поверхности они несут антигены и представляют их Т-лимфоцитам (хелперам).
Кроме того, в этой зоне находятся особые венулы, выстланные эндотелиоцитами кубической формы. Через стенку этих венул происходит переход Т- и В-лимфоцитов из крови в строму лимфатического узла.
Мозговое вещество лимфатических узлов является местом созревания плазматических клеток. Вместе со вторичными узелками коркового вещества мозговые тяжи составляют тимуснезависимую зону, или В-зону, лимфатических узлов. Мозговые тяжи кроме В-лимфоцитов и плазмоцитов содержат Т-лимфоциты и макрофаги.
Лимфа протекает через лимфатические узлы по синусам — пространствам, содержащим ретикулярную ткань, и ограниченным капсулой и трабекулами с одной стороны и узелками и мозговыми тяжами — с другой. Различают краевые, промежуточные и воротный синусы. Лимфа из последнего по лимфатическому сосуду выходит в области ворот. В просвете синусов обнаруживаются ретикулярные клетки, макрофаги, лимфоциты, плазматические клетки.
Возрастные изменения лимфатических узлов. С возрастом лимфатические узелки и их центры размножения постепенно исчезают, понижается фагоцитарная активность макрофагов, разрастается соединительная ткань трабекул, развиваются явления атрофии узлов и замещения их жировой тканью.
Реактивность и регенерация лимфатических узлов. Лимфатические узлы — весьма реактивные структуры. Они чувствительны к действию различных повреждающих факторов (радиации, инфекции, интоксикации и др.). Регенерация их возможна, если сохранены приносящие и выносящие лимфатические сосуды и, хотя бы частично, ретикулярная ткань, пролиферация клеток которой сопровождается заселением стволовыми клетками и последующей их дифференцировкой.
Учебное видео строения лимфатического узла
Скачать данное видео и просмотреть с другого видеохостинга можно на странице: Здесь.
– Также рекомендуем “Селезенка. Развитие селезенки. Строение селезенки.”
Оглавление темы “Выделительная система. Кроветворная система.”:
1. Плевра. Выделительный комплекс органов.
2. Почки. Строение почек. Нефрон. Функции и строение нефрона.
3. Петля Генле. Дистальный отдел нефрона. Собирательные трубочки почки. Сосуды почки.
4. Миоидные эндокриноциты. Юкставаскулярные клетки – Гурмагтига.
5. Мочевыводящие пути. Строение мочевыводящих путей.
6. Иммунный комплекс органов. Красный костный мозг.
7. Тимус. Развитие тимуса. Строение тимуса.
8. Лимфатические узлы. Развитие лимфатических узлов. Строение лимфатических узлов.
9. Селезенка. Развитие селезенки. Строение селезенки.
10. Иммунитет. Виды иммунитета. Виды иммунной реактивности организма.
Источник
Ретикуля́рная ткань (лат. textus reticularis) — особая форма соединительной ткани[1], состоящая из так называемых ретикулярных волокон и формирующих их ретикулярных клеток[en]. Ретикулярная ткань образует строму кроветворных органов и формирует микроокружение для клеток крови, формирующихся в них[2], однако обнаруживается и в органах, не связанных с кроветворением.
Строение[править | править код]
Ретикулярная ткань: схема строения (слева) и микрофотография (окрашивание солями серебра)
Ретикулярная ткань устроена как трёхмерная сеть, образованная стыкующимися ретикулярными фибробластоподобными[3]клетками, большинство из которых связаны с ретикулярными волокнами и окружают их своей цитоплазмой[4][2]. Благодаря отростчатой форме ретикулярные клетки выполняют механическую функцию. Они также секретируют компоненты основного вещества ретикулярной ткани — преколлаген, гликозаминогликаны, проэластин, микрофибриллярный белок[5].
Ретикулярные волокна достигают 0,5—2 мкм в диаметре и синтезируются ретикулярными клетками. Ретикулярные волокна содержат коллаген III типа и некоторых других типов, гликопротеины и протеогликаны. Они устойчивы к действию слабых кислот и щелочей, а также трипсина. Ретикулярные волокна являются аргирофильными, поскольку они выявляются при импрегнации солями серебра[en], которыми окрашиваются в чёрный цвет (при окрашивании гематоксилином и эозином они не видны). Аргирофильность ретикулярных волокон обусловлена с высоким содержанием углеводных цепей. К числу аргирофильных волокон также относят преколлагеновые волокна, которые представляют собой начальную форму образования коллагеновых волокон, однако ретикулярные волокна, по сравнению с преколлагеновыми, содержат серу, липиды и углеводы в высокой концентрации. В частности, ретикулярные волокна содержат 6—12 % гексоз против 1 % гексоз в коллагеновых волокнах (в отличие от ретикулярных, коллагеновые волокна содержат коллаген I типа[en], а не III типа). Ретикулярные волокна также красятся положительно при ШИК-реакции[en] за счёт углеводных цепей, как и при импреграции солями серебра. Из-за малого диаметра ретикулярные волокна при окрашивании сириусом красным[en] и визуализации с помощью поляризационной микроскопии[en]* приобретают зелёный цвет[6][7].
Ретикулярные волокна формируют тонкие, рыхло упакованные фибриллы, в которых отдельные волокна связаны друг с другом мостиками, сформированными протеогликанами и гликопротеинами. Фибриллы под электронным микроскопом имеют не всегда хорошо выраженную исчерченность с периодом от 64 до 67 нм. По растяжимости ретикулярные волокна занимают промежуточное положение между коллагеновыми и эластическими[6][7].
Функции[править | править код]
Ретикулярные волокна не являются уникальными только для ретикулярной ткани, но именно в ней являются доминирующими[8]. Ретикулярные волокна наиболее многочисленны в гладких мышцах, эндоневрии и строме кроветворных органов (селезёнки, лимфатических узлов, красного костного мозга), в которых они образуют ниши для развития клеток крови. Важную роль в формировании кроветворных ниш играют ретикулярные клетки, которые секретируют факторы роста, необходимые для определённых направлений развития гемопоэтических клеток[5]. Ретикулярные волокна также образуют сеть вокруг клеток некоторых паренхиматозных органов, таких как печень и эндокринные железы. За счет рыхлого расположения и малого диаметра ретикулярные волокна образуют гибкую сеть в органах, которые претерпевают изменение формы и объёма: артерии, селезёнка, печень, матка, мышцы, обеспечивающие перистальтику кишечника[9].
При генетически обусловленном дефиците коллагена III типа развивается синдром Элерса — Данлоса IV типа, при котором происходят разрывы структур, богатых ретикулярными волокнами, прежде всего артерий и кишечника[10].
Примечания[править | править код]
- ↑ Reticular tissue at Dorland’s Medical Dictionary (недоступная ссылка). Дата обращения: 16 июня 2009. Архивировано 16 июня 2009 года.
- ↑ 1 2 Афанасьев и др., 2004, с. 229.
- ↑ Жункейра, Карнейро, 2009, с. 302.
- ↑ Radivoj V. Krstic. Human Microscopic Anatomy: An Atlas for Students of Medicine and Biology (англ.). — Springer Science & Business Media, 2013. — P. 100. — ISBN 978-3-662-02676-2.
- ↑ 1 2 Афанасьев и др., 2004, с. 432.
- ↑ 1 2 Афанасьев и др., 2004, с. 229—230.
- ↑ 1 2 Жункейра, Карнейро, 2009, с. 132—134.
- ↑ Blue Histology – Connective Tissues. Дата обращения: 5 декабря 2008. Архивировано 16 мая 2013 года.
- ↑ Жункейра, Карнейро, 2009, с. 135—136.
- ↑ Жункейра, Карнейро, 2009, с. 136.
Литература[править | править код]
- Афанасьев Ю. И., Кузнецов С. Л., Юрина Н. А., Котовский Е. Ф. и др. Гистология, цитология и эмбриология. — 6-е изд., перераб. и доп.. — М.: Медицина, 2004. — 768 с. — ISBN 5-225-04858-7.
- Жункейра Л. К., Карнейро Ж. Гистология. — М.: ГЭОТАР-Медиа, 2009. — 576 с. — ISBN 978-5-9704-1352-4.
Источник
Ретикулярная ткань (textus connectivus reticularis (LNH); лат. reticulum сетка; син. сетчатая ткань) — разновидность соединительной ткани, состоящей из ретикулярных клеток и ретикулярных волокон, заключенных в основное межклеточное вещество и образующих рыхлую трехмерную сеть, являющуюся основой кроветворных и лимфоидных органов.
Представления о Ретикулярной ткани формировались в течение многих десятилетий, но лишь в последние 10—15 лет, благодаря использованию достижений цитогенетики, радиобиологии, иммунологии, трансплантологии, были получены принципиально новые данные о ее дифференцировке, структуре и функции.
В эмбриогенезе Ретикулярной ткани дифференцируется из мезенхимы (см.) и в раннем постнатальном периоде постепенно приобретает строение зрелой ткани.

Рис. 1. Электронограмма ретикулярной клетки костного мозга мыши: 1 — ядро; 2 — цитоплазматические отростки; 3 — зернистая эндоплазматическая сеть; X 5000.
Ретикулярные клетки (cellulae reticulares) относят к клеткам фибробластического типа, наряду с фибробластами, хондробластами и остеобластами, объединяемыми под названием «механоциты». Ретикулярные клетки (рис. 1) имеют уплощенную, веретеновидную или звездчатую форму с гладкой поверхностью; ядро — угловатое или вытянутое. Степень развития органелл, в частности зернистой эндоплазматической сети и комплекса Гольджи (см. Гольджи комплекс), вариабельна и зависит от функционального состояния ретикулярной клетки. Соседние ретикулярные клетки или отростки одних и тех же клеток контактируют друг с другом посредством соединений типа промежуточных или десмосом (см.).
Ввиду того, что ретикулярные клетки часто трудно различить среди массы кроветворных клеток, их идентификация возможна только при использовании электронной микроскопии (см.), гистохимических методов исследования (см.) и методов иммуногистохимии (см. Иммуноморфология) или комбинации этих методов, позволяющих с высокой степенью достоверности выявлять ретикулярные клетки, а также идентифицировать их от внешне сходных с ними клеток системы мононуклеарных фагоцитов (см.).
Гистохимические свойства ретикулярных клеток определяются органными, а также видовыми особенностями. У человека ретикулярные клетки белой пульпы селезенки (см.) отличаются от макрофагов (см.) более низкой активностью эстеразы (см.) и кислой фосфатазы (см.), а от интердигитирующих клеток (разновидности мононуклеарных фагоцитов) — отсутствием активности АТФ-азы. В ретикулярных клетках лимфоцитарной короны лимф, фолликулов селезенки выявляется отчетливая активность 5′-нуклеотидазы (табл.).
Активность 5′-нуклеотидазы определяется также в ретикулярных клетках белой пульпы селезенки крыс и морских свинок, но отсутствует у кроликов. Ретикулярные клетки костного мозга мышей и крыс характеризуются активностью щелочной фосфатазы, локализующейся на их плазматической мембране. Этим признаком они отличаются от макрофагов, в к-рых выявляется активность кислой фосфатазы, локализующейся в лизосомах (см.). Такой морфол. тип ретикулярных клеток часто называют фибробластическим.
Рис. 2. Схематическое изображение участка лимфоидного фолликула (взаиморасположение дендритной ретикулярной клетки и окружающих образований): 1 — ядро дендрит ной ретикулярной клетки с ядрышком (окрашено в темный цвет); 2 — митохондрия; 3 — гранулярная эндоплазматическая сеть; 4 — агранулярная эндоплазматическая сеть; 5 — отростки дендритной ретикулярной клетки; 6 — лимфоциты; 7 — десмосомы; 8 — капилляр.
В светлых (герминативных) центрах лимфатических фолликулов селезенки и лимфатических узлов (см.) описан особый тип ретикулярных клеток — дендритные ретикулярные клетки. Их отличают крупные размеры, ядро неправильной формы с выраженным ядрышком, многочисленные гладкие пузырьки в цитоплазме. Характерным признаком дендритных ретикулярных клеток являются длинные ветвящиеся отростки с многочисленными складками, глубоко проникающие между окружающими лимфоидными клетками (рис. 2). Отростки дендритных ретикулярных клеток вместе с цитоплазматическими выпячиваниями иммунобластов (В-лимфоцитов) формируют сложную сеть в виде лабиринта. В межклеточном пространстве среди отростков локализуются микровезикулярные структуры и глобулярные плотные частицы диаметром 20—70 нм. В области контакта отростков дендритных ретикулярных клеток видны структуры типа десмосом (см.). В отличие от макрофагов с окрашивающимися включениями и других мононуклеарных фагоцитов дендритные ретикулярные клетки имеют более низкую активность эстеразы и кислой фосфатазы, но высокую активность 5′-нуклеотидазы. По ряду гистохимических признаков они могут быть сходны с мононуклеарными фагоцитами: макрофагами красной пульпы селезенки, металлофильными (аргирофильными) клетками краевой зоны лимф, фолликулов, макрофагами с окрашивающимися включениями. Решающим критерием для идентификации дендритных ретикулярных клеток является способность этих клеток связывать (но не фагоцитировать) иммунные комплексы на своей поверхности.
Ретикулярные волокна (fibrae reticulares), входящие в состав Р. т., состоят из фибрилл различного диаметра, заключенных в гомогенное, плотное основное межклеточное вещество. Фибриллы диаметром 20—50 нм имеют осевую исчерченность, характерную для зрелого коллагена. Тонкие фибриллы диаметром 10 нм локализуются по периферии ретикулярных волокон. Ретикулярные волокна всегда окутаны цитоплазмой ретикулярных клеток в виде чехла (инвагинированы в цитолемму), просвет к-рого всегда сообщается с межклеточным пространством. В лимфоидных органах сеть ретикулярных волокон развита сильнее, чем в костном мозге, причем в тимус-зависимых зонах лимфатических узлов она более рыхлая, чем в корковом и мозговом веществе.
По данным биохимического и иммунохимического анализов, ретикулярные волокна состоят из коллагена III типа (см. Коллаген) и неколлагенового компонента в виде аморфного межфибриллярного вещества с выраженными иммуногенными свойствами. В составе неколлагенового компонента идентифицированы белки (90%), углеводы (4%) и липиды (4%). Для выявления ретикулярных волокон широко используют различные варианты импрегнации серебром. В повседневную практику начинают входить иммуногистохимические и иммунофлюоресцентные методы выявления ретикулярных волокон (см. Иммуноморфология, Иммунофлюоресценция), в основе к-рых лежит использование специфических антисывороток к коллагенам разных типов.
Р. т. образует строму и является носителем специфических органных функций костного мозга и периферических лимфоидных органов (селезенки, лимф, узлов, солитарных и групповых лимф, фолликулов жел.-киш. тракта). После того как было экспериментально доказано, что ретикулярные клетки гистогенетически независимы от кроветворных, их стали относить к категории стромальных элементов, ответственных за создание специфического микроокружения, обеспечивающего миграцию, сортировку, репликацию и дифференцировку кроветворных и лимфоидных клеток. Ретикулярные клетки способны восстанавливать исходное микроокружение при повреждении органов, содержащих Р. т., или переносить его при эктопической трансплантации. Во всех случаях сначала восстанавливается ретикулярная строма, а затем она репопулируется (заселяется) кроветворными или лимфоидными клетками (см. Лимфоидная ткань). В костном мозге ретикулярные клетки формируют также адвентициальный слой синусоидных капилляров и в условиях нормального кроветворения покрывают до 60% их поверхности. Длинные ветвящиеся отростки ретикулярных клеток вступают в специфическое взаимодействие с кроветворными клетками гранулоцитарно-го ряда дифференцировки (см. Кроветворные органы). Концентрация ретикулярных клеток повышена вблизи эндоста.
Экспериментальные данные свидетельствуют о сохранении в пост-натальном онтогенезе гистогенети-ческой близости между разными типами механоцитов (фибробластами, хрящевыми, костными, ретикулярными клетками). Так, при культивировании костного мозга in vitro возникают колонии-клоны фибробластов; нек-рые из этих колоний при обратной трансплантации в организм формируют костномозговой орган (участок костной ткани, окружающей костный мозг), заселяемый кроветворными клетками. Эти данные показывают, что среди ретикулярных клеток костного мозга имеются элементы, способные трансформироваться в истинные фибробласты (судя по их способности синтезировать коллаген I и III типов), и в то же время, проявляющие остеогенные свойства. В другой экспериментальной модели костномозговой орган возникает в результате последовательных морфогенетических процессов под влиянием индуктора — деминерализованного костного матрикса, имплантированного в подкожную соединительную ткань. По мнению Редди, Гея, Гея, Миллера (А. N. Beddi, В. Gay, S. Gay. E. J. Miller, 1977), в этом случае под влиянием индуктора происходит последовательная трансформация фибробластов в хрящевые, костные, а затем ретикулярные (стромальные) клетки. По-видимому, ретикулярные клетки костного мозга способны также трансформироваться в жировые клетки, участвующие в создании кроветворного микроокружения (в красном костном мозге). В норме ретикулярные клетки отличает высокая радиорезистентность, и они практически не делятся.
Существует мнение, что на поверхности дендритных ретикулярных клеток имеются мембранные рецепторы для иммуноглобулинов (см.), с к-рыми связываются иммунные комплексы антигенов с антителами. Согласно другому мнению, антигены (см.) просто медленно фильтруются через лабиринт, создаваемый отростками дендритных ретикулярных клеток. Связывание антигена происходит быстрее у предварительно иммунизированных животных (через 0,5—2 часа), чем у неиммунизированных (через 4—24 часа). Параллельно изменяется и ультраструктура дендритных ретикулярных клеток. В опытах на кроликах показано, что в процессе формирования светлых центров дендритные ретикулярные клетки трансформируются из фибробластических ретикулярных клеток прилежащей лимфоцитарной короны. При этом ретикулярные клетки теряют активность щелочной фосфатазы (и, по-видимому, способность к волокнообразованию).
С возрастом в Ретикулярной ткани увеличивается количество волокнистых структур. Для костного мозга характерно необратимое замещение ретикулярной стромы жировой тканью и прекращение кроветворения. С возрастными и патологическими изменениями Р. т. тесно связано нарушение функции кроветворных и лимфоидных органов. При воздействии высоких доз ионизирующего излучения на организм Р. т. не регенерирует, а замещается фиброзной. Миелофиброз характеризуется интенсивной пролиферацией фиброгенных клеток и последующим массивным отложением коллагена I, III и IV типов. Показано участие ретикулярных клеток в синтезе фибриллярных белков амилоида (см. Амилоидоз). Поражение собственно ретикулярной стромы, сопровождающееся нарушением кроветворения, не всегда можно выявить морфологически. Поэтому важная роль отводится методам клонирования стромальных клеток in vitro, изучению их взаимодействия с кроветворными и лимфоидными клетками в жидкостных и агаровых культурах.
См. также Соединительная ткань.
Таблица. Сравнительная характеристика активности ферментов в ретикулярных клетках и мононуклеарных фагоцитах (макрофагах) белой пульпы селезенки человека По данным Мюллер-Хермелинка (H. К. Muller-Hermelink) и др. (1974)
Название фермента | Активность ферментов в различных клетках лимфатического | |||
Ретикулярные клетки лимфоцитарной короны | Дендритные ретикулярные клетки светлых центров | Макрофаги | Интердигитирующие клетки тимусзависимой зоны | |
Эстераза | + | + | ++ | + |
Кислая фосфатаза | +/- | +/- | ++ | + |
АТФ-аза | – | – | +/- | ++ |
5′-нуклеотидаза | + | ++ | – | – |
Щелочная фосфатаза | – | – | – | – |
где ++ очень высокая + высокая +/- низкая – отсутствует | ||||
Библиография: Старостин В. И. и Мичурина Т. В. Строма кроветворных органов и ее взаимоотношение со стволовой кроветворной клеткой, в кн.: Морфол. человека и животных, Антропология, под ред. Н. Г. Хрущова и В. И. Старостина, с. 59, М., 1977, библиогр.; Фриденштейн А. Я. и Лурия Е. А. Клеточные основы кроветворного микроокружения, М., 1980, библиогр.; Castro-Mаlaspinа Н. а. о. Characterization of human bone marrow fibroblast colony-forming cells (CFU-F) and their progeny, Blood, v. 56, p. 289, 1980; Duсastelle Т., Rey nes M. et Die-bold J. Les cellules non lymphoides des organes lymphoides рёпрЬёпдиез, Path, et Biol., t. 29, p. Ill, 1981, bibliogr.; Gay S. a. Miller E. J. Collagen in the physiology and pathology of connective tissue, Stuttgart — N. Y., 1978; Groscurth P. Non-lymphatic cells in the lymph node cortex of the mouse, Path, res. Pract., v. 169, p. 212, 235, 255, 1980; Mononuclear phagocytes, In immunity, infection and pathology, ed. by R. van Furth, Oxford a. o., 1975; Mononuclear phagocytes, Functional aspects, ed. by R. van Furth, pt 1—2, Hague a. o., 1980; Puchtler H. a. Waldrop F. S. Silver impregnation methods for reticulum filers and reticulin, a reinvestigation of their origins and specifity, Histochemistry, v. 57, p. 177, 1978; Westen H. a. Bainton D. F. Association of alkaline-phosphatase-positive reticulum cells in bone marrow with granulocytic precursors, J. exp. Med., v. 150, p. 919, 1979.
H. Г. Хрущов, В. И. Старостин.
Источник