Реакция т и в лимфоцитов на антиген

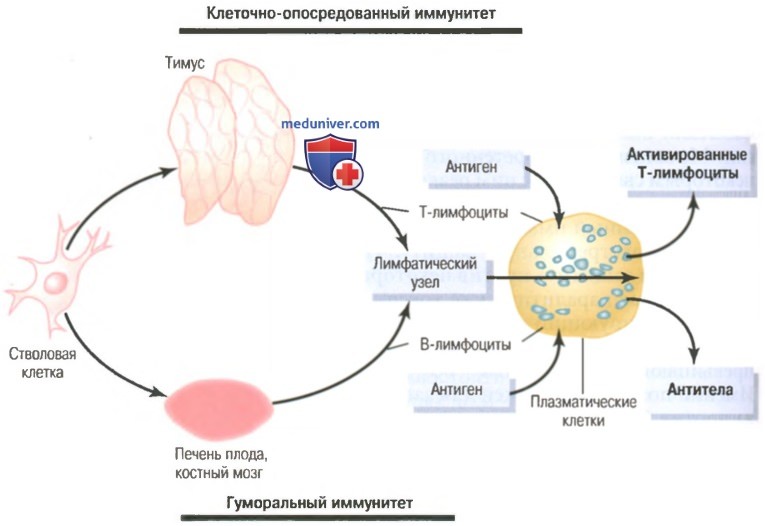
Т-лимфоциты и В-лимфоциты иммунитета. Предварительная обработка Т- и В-лимфоцитова) Клеточно-опосредованный и гуморальный иммунитеты обеспечивают два типа лимфоцитов – Т- и В-лимфоциты. Хотя при исследовании под микроскопом большинство лимфоцитов в нормальной лимфоидной ткани выглядят одинаковыми, эти клетки подразделяют на две основные популяции. Одна популяция — Т-лимфоциты — ответственна за формирование активированных лимфоцитов, обеспечивающих клеточно-опосредованный иммунитет. Другая популяция — В-лимфоциты — ответственна за формирование антител, обеспечивающих гуморальный иммунитет. Оба типа лимфоцитов образуются у эмбриона из полипотентных гемопоэтических стволовых клеток, формирующих лимфоциты как один из наиболее важных результатов их дифференцировки. Почти все сформированные лимфоциты в результате заселяют лимфоидную ткань, однако прежде чем это произойдет, они дополнительно дифференцируются или проходят предварительную обработку. Лимфоциты, которые в итоге станут активированными Т-лимфоцитами, сначала мигрируют к тимусу, где подвергаются предварительной обработке. Эти ответственные за клеточно-опо-средованный иммунитет лимфоциты называют Т-лимфоцитами, что подчеркивает роль тимуса. Другая популяция лимфоцитов, В-лимфоциты, предназначенные для формирования антител, проходят предварительную обработку в печени плода в середине периода внутриутробной жизни, а также в костном мозге в конце внутриутробной жизни плода и после рождения. Эта популяция клеток впервые была открыта у птиц, имеющих специальный орган для их предварительной обработки, который называют бурсой Фабриция (фабрициевой сумкой). Ответственные за гуморальный иммунитет лимфоциты называют В-лимфоцитами, что подчеркивает роль бурсы. На рисунке ниже показаны две лимфоцитарные системы для формирования: (1) активированных Т-лимфоцитов; (2) антител.
Предварительная обработка Т- и В-лимфоцитовВсе лимфоциты организма происходят от коммитированных в лимфоцитарном направлении стволовых клеток эмбриона, но эти клетки не могут непосредственно превратиться в активированные Т-лимфоциты или антитела. Прежде чем это станет возможным, клетки должны подвергнуться дальнейшей дифференцировке в соответствующих областях, где они проходят специфическую обработку. а) Т-лимфоциты проходят предварительную обработку в тимусе (вилочковой железе). После образования в костном мозге Т-лимфоциты сначала мигрируют к вилочковой железе. Здесь они быстро делятся, одновременно становясь чрезвычайно разнообразными, т.е. предназначенными для реакции против разных специфических антигенов. Это значит, что один лимфоцит, обработанный в тимусе, проявляет специфическую реактивность в отношении одного антигена. Следующий лимфоцит специфически реагирует на другой антиген. Это продолжается до тех пор, пока в тимусе не появятся тысячи разных типов лимфоцитов со специфической реактивностью в отношении тысяч разных антигенов. Эти разные типы предварительно обработанных Т-лимфоцитов оставляют тимус и распространяются кровью по всему телу, временно оседая в лимфоидной ткани. Кроме того, благодаря обработке в тимусе любой оставляющий его Т-лимфоцит не реагирует с белками или другими антигенами собственных тканей организма (иначе Т-лимфоциты погубили бы собственное тело человека в течение всего нескольких дней). Тимус выбирает, какие Т-лимфоциты могут его покинуть, сначала смешивая их практически со всеми специфическими аутоантигенами собственных тканей тела. Если Т-лимфоцит реагирует, он разрушается и фагоцитируется, вместо того, чтобы выделяться. Это происходит с основной частью клеток (вплоть до 90%). Таким образом, клетки, выделяющиеся из тимуса, не реагируют против собственных антигенов тела; они реагируют лишь на антигены внешних источников, например бактерий, токсинов или тканей, пересаженных от другого человека. Основная часть предобработки Т-лимфоцитов в тимусе происходит перед рождением ребенка и в течение нескольких месяцев после рождения. Удаление вилочковой железы после этого периода ослабляет (но не исключает) Т-лимфоцитарную иммунную систему. Однако удаление тимуса за несколько месяцев до рождения может нарушить развитие всего клеточно-опосредован-ного иммунитета. Поскольку именно клеточный тип иммунитета в основном отвечает за отторжение трансплантированных органов, например сердца или почек, органы можно пересаживать с меньшей вероятностью отторжения, если у животного в соответствующее время до его рождения удалить тимус. б) В-лимфоциты проходят предварительную обработку в печени и костном мозге. О деталях предварительной обработки В-лимфоцитов известно гораздо меньше, чем о предобработке Т-лимфоцитов. Известно, что у человека предварительная обработка В-лимфоцитов осуществляется в печени в середине внутриутробного периода развития, а также в костном мозге в конце внутриутробного периода и после рождения. Существуют два важных различия между В- и Т-лимфоцитами. Во-первых, В-лимфоциты активно секретируют реактивные агенты, называемые антителами, в отличие от Т-лимфоцитов, реагирующих с антигеном непосредственно. Антитела — это крупные белковые молекулы, способные соединяться с антигенной субстанцией и разрушать ее. Во-вторых, разнообразие В-лимфоцитов выражено больше, чем у Т-лимфоцитов, т.е. формируются миллионы типов В-лимфоцитарных антител с разными специфическими реактивностями. После предобработки В-лимфоциты, как и Т-лимфоциты, мигрируют к лимфоидной ткани по всему телу, где временно располагаются рядом, но несколько обособленно от областей локализации Т-лимфоцитов. – Также рекомендуем “Роль лимфоцитарных клонов. Происхождение клонов лимфоцитов” Оглавление темы “Клетки иммунитета. Виды иммунитета”: |
Источник
Активация В-лимфоцитов. Влияние антигенов на В-лимфоцитыАктивация В-лимфоцитов связана с рядом специфических и неспецифических стимулов, которые приводят к глубоким биохимическим и морфологическим изменениям в этих клетках. Существуют теории, пытающиеся объяснить, каким образом антиген «запускает», активирует В-клетки. Хотя действие антигена и является наиболее важным для активации В-клеток, в процессе активации принимают участие и дополнительные факторы: 1) антигенспецифические Т-клетки, 2) медиаторы, секретируемые антигенспецифическими Т-клетками, 3) медиаторы, секретируемые неспецифическими Т-клетками, 4) продукт 1а-гена, 5) добавочные клетки (макрофаги, А-клетки), 6) антигенспецифические Т-супрессоры. 1. Активация В-клеток обусловлена внедрением липофильного блока в двойной липидный слой их мембраны. Это внедрение может происходить и без участия антигена, неспецифически, при большой концентрации соответствующего метаболита. Таким образом, например, действует, по-видимому, неспецифический стимулирующий фактор Т-клеток-помощников. Специфичность иммунологического процесса обусловлена наличием медиатора, который секретируется Т-лимфоцитами под влиянием антигена. Этот медиатор (секретируемый рецептор Т-клеток) состоит из трех частей: рецептора Т-клеток, специфически реагирующего с детерминантами несущей части антигена (носителя), Н-2-белка и «липофильного хвоста». Медиатор присоединяется своим рецептором к антигену, фиксированному на поверхности антигенсвязывающей клетки, и липофильный хвост внедряется в фосфолипидный слой мембраны.
2. Присоединение антигена к В-лимфоциту приводит к его обратимому параличу или инактивации благодаря возникновению сигнала 1. Сигнал 1 развивается при бимолекулярной реакции между одновалентным антигеном и соответствующим рецептором В-клетки. Он чрезвычайно быстро (возможно, уже через секунду) оказывает на клетку парализующее действие. Индукция иммунной реакции возникает в том случае, если на В-клетку после сигнала 1 подействует еще и сигнал 2. Первоначально предполагалось, что сигнал 2 вызывают антитела, которые фиксированы на поверхности Т-клеток и специфичны для антигена, присоединившегося к В-лимфоцитам. В дальнейшем была допущена возможность передачи сигнала 2 на небольшое расстояние при помощи образуемого Т-лимфоцитами медиатора. В некоторых случаях сигнал 2 обусловлен действием антител не против антигенов, присоединившихся к поверхности В-лимфоцитов, а против собственных антигенов его поверхности. 3. Для активации В-лимфоцитов необходим второй сигнал. Этот сигнал исходит от присоединившегося к клетке С3-компонента комплемента. В пользу этого взгляда говорят наличие на поверхности ПАОК рецепторов для активизированного С3-компонента и митогенность очищенных препаратов С3-компонента. Соответствует этой точке зрения и наличие в лимфатических клетках липосомальных протеаз, активирующих С3-компонент, и то, что эти протеазы выделяются как при действии на лимфатические клетки антигенов и иммуногенов, так и при взаимодействии Т- и В-лимфоцитов (Dukor, Hartman, 1973; Hartman, 1975). В связи с этой точкой зрения нельзя не упомянуть о митогенном действии протеаз. Действие на клетки селезенки трипсина стимулирует включение 3Н-тимидина в эти клетки почти так же сильно, как действие самых мощных митогенов. Действие трипсина направлено на В-клетки: более 80% трансформированных им клеток содержало на поверхности иммуноглобулины; он чрезвычайно сильно стимулирует включение 3Н-тимидина в клетки селезенок бестимусных мышей (Kaplan, Bona, 1974; Vischer, 1974). 4. По модели одного неспецифического сигнала присоединение антигена к иммуноглобулиновым рецепторам В-лимфоцитов индуцирует ряд процессов (например, образование «колпачка»), но не активирует эти клетки. В-лимфоциты активируются неспецифическим сигналом от участка на их поверхности, который не является иммуноглобулином. Эти сигналы исходят от самого антигена (но не от их антигенной детерминанты) в случае тимуснезависимых антигенов и от медиаторов Т-клеток или макрофагов (в случае тимусзависимых антигенов). – Также рекомендуем “Результаты активации В-лимфоцитов. Изменения В-клеток после активации” Оглавление темы “Антителообразующие клетки. Контроль образования антител”: |
Источник
Оглавление темы “Селезенка. Лимфатические узлы. Стадии иммунного ответа. Формы иммунного ответа. Воспаление. Механизмы, контролирующие иммунную систему.”: Активация Т- и В-лимфоцитов в иммунном ответе. Активация лимфоцитов. Формы специфического иммунного ответа.Передача сигнала активации от антигенраспознающего рецептора в ядро клетки приводит к активации конкретных генов, ответственных за митотическую активность клеток, контролирующих продукцию клеткой биологически активных молекул, участвующих в антимикробной защите (антитела, цитокины).
В-лимфоцит получает одновременно два сигнала активации: от антигенраспознающего рецептора при его соединении с антигеном и от связывания его поверхностных костимулирующих молекул с соответствующими лигандами на Т-лимфоцитах. После этого В-лимфоцит пролиферирует и потомки его превращаются в зрелые антителопродуцирующие плазматические клетки. Т-лимфоцит в ответ на контакт с антигеном начинает пролиферировать, потомки его приобретают способность продуцировать определенные цитокины или превращаются в зрелые цитотоксические клетки. В зависимости от того, какие дополнительные сигналы активации (цитокины, костимулирующие молекулы) получает Т-лимфоцит в момент контакта с антигеном, его потомки дифференцируются в двух разных направлениях, превращаясь в Т-хелперы, продуцирующие гамма-интерферон (Th1), или в Т-хелперы, продуцирующие интерлейкины-4, -5, -6, -10, -13 (Th2). Количественное преобладание Thl над Th2 является условием развития клеточного (кле-точно-опосредованного) иммунного ответа. В случае преобладания Th2 развивается гуморальный иммунный ответ, проявляющийся продукцией специфических антител (рис. 8.9). – Также рекомендуем “Клеточный иммунный ответ. Гуморальный иммунный ответ. Защитные функции иммуноглобулинов ( антител ).” |
Источник
№ 10 Иммунокомпетентные клетки. Т- и В-лимфоциты, макрофаги, их кооперация.
Иммунокомпетентные клетки – клетки, способные специфически распознавать антиген и отвечать на него иммунной реакцией. Такими клетками являются Т- и В-лимфоциты (тимусзависимые и костномозговые лимфоциты), которые под влиянием чужеродных агентов дифференцируются в сенсибилизированный лимфоцит и плазматическую клетку.
Т-лимфоциты – это сложная по составу группа клеток, которая происходит от полипотентной стволовой клетки костного мозга, а созревает и дифференцируется в тимусе из предшественников. Т-лимфоциты разделяются на две субпопуляции: иммунорегуляторы и эффекторы. Задачу регуляции иммунного ответа выполняют Т-хелперы. Эффекторную функцию осуществляют Т-киллеры и естественные киллеры. В организме Т-лимфоциты обеспечивают клеточные формы иммунного ответа, определяют силу и продолжительность иммунной реакции.
B-лимфоциты – преимущественно эффекторные иммунокомпетентные клетки. Зрелые В-лимфоциты и их потомки – плазматические клетки являются антителопродуцентами. Их основными продуктами являются иммуноглобулины. В-лимфоциты участвуют в формировании гуморального иммунитета, В-клеточной иммунологической памяти и гиперчувствительности немедленного типа.
Макрофаги – клетки соединительной ткани, способные к активному захвату и перевариванию бактерий, остатков клеток и других, чужеродных для организма частиц. Основная функция макрофагов сводится к борьбе с теми бактериями, вирусами и простейшими, которые могут существовать внутри клетки-хозяина, при помощи мощных бактерицидных механизмов. Роль макрофагов в иммунитете исключительно важна – они обеспечивают фагоцитоз, переработку и представление антигена T-клеткам.
Кооперация иммунокомпетентных клеток. Иммунная реакция организма может иметь различный характер, но всегда начинается с захвата антигена макрофагами крови и тканей или же со связывания со стромой лимфоидных органов. Нередко антиген адсорбируется также на клетках паренхиматозных органов. В макрофагах он может полностью разрушаться, но чаще подвергается лишь частичной деградации. В частности, большинство антигенов в лизосомах фагоцитов в течение часа подвергается ограниченной денатурации и протеолизу. Оставшиеся от них пептиды (как правило, два-три остатка аминокислот) комплексируются с экспрессированными на внешней мембране макрофагов молекулами МНС.
Макрофаги и все другие вспомогательные клетки, несущие на внешней мембране антигены, называются антигенпрезентирующими, именно благодаря им Т- и В-лимфоциты, выполняя функцию презентации, позволяют быстро распознавать антиген.
Иммунный ответ в виде антителообразования происходит при распознавании В-клетками антигена, который индуцирует их пролиферацию и дифференциацию в плазмоцит. Прямое воздействие на В-клетку без участия Т-клеток могут оказать только тимуснезависимые антигены. В этом случае В-клетки кооперируются с Т-хелперами и макрофагами. Кооперация на тимусзависимый антиген начинается с его презентации на макрофаге Т-хелперу. В механизме этого распознавания ключевую роль имеют молекулы МНС, так как рецепторы Т-хелперов распознают номинальный антиген как комплекс в целом или же, как модифицированные номинальным антигеном молекулы МНС, приобретшие чужеродность. Распознав антиген, Т-хелперы секретируют γ-интерферон, который активирует макрофаги и способствует уничтожению захваченных ими микроорганизмов. Хелперный эффект на В-клетки проявляется пролиферацией и дифференциацией их в плазмоциты. В распознавании антигена при клеточном характере иммунного ответа, кроме Т-хелперов, участвуют также Т-киллеры, которые обнаруживают антиген на тех антигенпрезентирующих клетках, где он комплексируется с молекулами МНС. Более того, Т-киллеры, обусловливающие цитолиз, способны распознавать не только трансформированный, но и нативный антиген. Приобретая способность вызывать цитолиз, Т-киллеры связываются с комплексом антиген + молекулы МНС класса 1 на клетках-мишенях; привлекают к месту соприкосновения с ними цитоплазматические гранулы; повреждают мембраны мишеней после экзоцитоза их содержимого.
В результате продуцируемые Т-киллерами лимфотоксины вызывают гибель всех трансформированных клеток организма, причем особенно чувствительны к нему клетки, зараженные вирусом. При этом наряду с лимфотоксином активированные Т-киллеры синтезируют интерферон, который препятствует проникновению вирусов в окружающие клетки и индуцирует в клетках образование рецепторов лимфотоксина, тем самым повышая их чувствительность к литическому действию Т-киллеров.
Кооперируясь в распознавании и элиминации антигенов, Т-хелперы и Т-киллеры не только активируют друг друга и своих предшественников, но и макрофагов. Те же, в свою очередь, стимулируют активность различных субпопуляций лимфоцитов.
Регуляция клеточного иммунного ответа, как и гуморального, осуществляется Т-супрессорами, которые воздействуют на пролиферацию цитотоксических и антигенпрезентирующих клеток.
Цитокины. Все процессы кооперативных взаимодействий иммунокомпетентных клеток, независимо от характера иммунного ответа, обусловливаются особыми веществами с медиаторными свойствами, которые секретируются Т-хелперами, Т-киллерами, мононуклеарными фагоцитами и некоторыми другими клетками, участвующими в реализации клеточного иммунитета. Все их многообразие принято называть цитокинами. По структуре цитокины являются протеинами, а по эффекту действия — медиаторами. Вырабатываются они при иммунных реакциях и обладают потенциирующим и аддитивным действием; быстро синтезируясь, цитокины расходуются в короткие сроки. При угасании иммунной реакции синтез цитокинов прекращается.
Источник