Образование системы т и в лимфоцитов

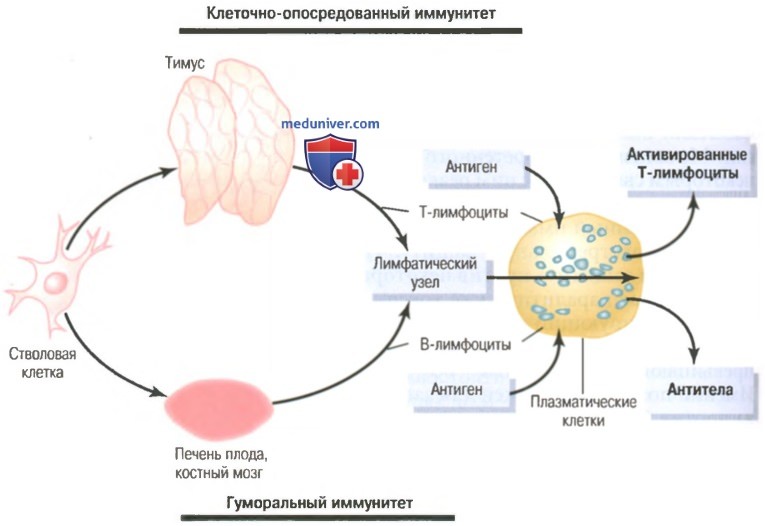
Т-лимфоциты и В-лимфоциты иммунитета. Предварительная обработка Т- и В-лимфоцитова) Клеточно-опосредованный и гуморальный иммунитеты обеспечивают два типа лимфоцитов – Т- и В-лимфоциты. Хотя при исследовании под микроскопом большинство лимфоцитов в нормальной лимфоидной ткани выглядят одинаковыми, эти клетки подразделяют на две основные популяции. Одна популяция — Т-лимфоциты — ответственна за формирование активированных лимфоцитов, обеспечивающих клеточно-опосредованный иммунитет. Другая популяция — В-лимфоциты — ответственна за формирование антител, обеспечивающих гуморальный иммунитет. Оба типа лимфоцитов образуются у эмбриона из полипотентных гемопоэтических стволовых клеток, формирующих лимфоциты как один из наиболее важных результатов их дифференцировки. Почти все сформированные лимфоциты в результате заселяют лимфоидную ткань, однако прежде чем это произойдет, они дополнительно дифференцируются или проходят предварительную обработку. Лимфоциты, которые в итоге станут активированными Т-лимфоцитами, сначала мигрируют к тимусу, где подвергаются предварительной обработке. Эти ответственные за клеточно-опо-средованный иммунитет лимфоциты называют Т-лимфоцитами, что подчеркивает роль тимуса. Другая популяция лимфоцитов, В-лимфоциты, предназначенные для формирования антител, проходят предварительную обработку в печени плода в середине периода внутриутробной жизни, а также в костном мозге в конце внутриутробной жизни плода и после рождения. Эта популяция клеток впервые была открыта у птиц, имеющих специальный орган для их предварительной обработки, который называют бурсой Фабриция (фабрициевой сумкой). Ответственные за гуморальный иммунитет лимфоциты называют В-лимфоцитами, что подчеркивает роль бурсы. На рисунке ниже показаны две лимфоцитарные системы для формирования: (1) активированных Т-лимфоцитов; (2) антител.
Предварительная обработка Т- и В-лимфоцитовВсе лимфоциты организма происходят от коммитированных в лимфоцитарном направлении стволовых клеток эмбриона, но эти клетки не могут непосредственно превратиться в активированные Т-лимфоциты или антитела. Прежде чем это станет возможным, клетки должны подвергнуться дальнейшей дифференцировке в соответствующих областях, где они проходят специфическую обработку. а) Т-лимфоциты проходят предварительную обработку в тимусе (вилочковой железе). После образования в костном мозге Т-лимфоциты сначала мигрируют к вилочковой железе. Здесь они быстро делятся, одновременно становясь чрезвычайно разнообразными, т.е. предназначенными для реакции против разных специфических антигенов. Это значит, что один лимфоцит, обработанный в тимусе, проявляет специфическую реактивность в отношении одного антигена. Следующий лимфоцит специфически реагирует на другой антиген. Это продолжается до тех пор, пока в тимусе не появятся тысячи разных типов лимфоцитов со специфической реактивностью в отношении тысяч разных антигенов. Эти разные типы предварительно обработанных Т-лимфоцитов оставляют тимус и распространяются кровью по всему телу, временно оседая в лимфоидной ткани. Кроме того, благодаря обработке в тимусе любой оставляющий его Т-лимфоцит не реагирует с белками или другими антигенами собственных тканей организма (иначе Т-лимфоциты погубили бы собственное тело человека в течение всего нескольких дней). Тимус выбирает, какие Т-лимфоциты могут его покинуть, сначала смешивая их практически со всеми специфическими аутоантигенами собственных тканей тела. Если Т-лимфоцит реагирует, он разрушается и фагоцитируется, вместо того, чтобы выделяться. Это происходит с основной частью клеток (вплоть до 90%). Таким образом, клетки, выделяющиеся из тимуса, не реагируют против собственных антигенов тела; они реагируют лишь на антигены внешних источников, например бактерий, токсинов или тканей, пересаженных от другого человека. Основная часть предобработки Т-лимфоцитов в тимусе происходит перед рождением ребенка и в течение нескольких месяцев после рождения. Удаление вилочковой железы после этого периода ослабляет (но не исключает) Т-лимфоцитарную иммунную систему. Однако удаление тимуса за несколько месяцев до рождения может нарушить развитие всего клеточно-опосредован-ного иммунитета. Поскольку именно клеточный тип иммунитета в основном отвечает за отторжение трансплантированных органов, например сердца или почек, органы можно пересаживать с меньшей вероятностью отторжения, если у животного в соответствующее время до его рождения удалить тимус. б) В-лимфоциты проходят предварительную обработку в печени и костном мозге. О деталях предварительной обработки В-лимфоцитов известно гораздо меньше, чем о предобработке Т-лимфоцитов. Известно, что у человека предварительная обработка В-лимфоцитов осуществляется в печени в середине внутриутробного периода развития, а также в костном мозге в конце внутриутробного периода и после рождения. Существуют два важных различия между В- и Т-лимфоцитами. Во-первых, В-лимфоциты активно секретируют реактивные агенты, называемые антителами, в отличие от Т-лимфоцитов, реагирующих с антигеном непосредственно. Антитела — это крупные белковые молекулы, способные соединяться с антигенной субстанцией и разрушать ее. Во-вторых, разнообразие В-лимфоцитов выражено больше, чем у Т-лимфоцитов, т.е. формируются миллионы типов В-лимфоцитарных антител с разными специфическими реактивностями. После предобработки В-лимфоциты, как и Т-лимфоциты, мигрируют к лимфоидной ткани по всему телу, где временно располагаются рядом, но несколько обособленно от областей локализации Т-лимфоцитов. – Также рекомендуем “Роль лимфоцитарных клонов. Происхождение клонов лимфоцитов” Оглавление темы “Клетки иммунитета. Виды иммунитета”: |
Источник
Оглавление темы “Иммунная система. Функции клеток иммунной системы. Т-лимфоциты. В-лимфоциты. Структура и функции органов иммунной системы.”:
1. Иммунная система. Иммунная система человека. Неспецифический иммунитет. Специфический иммунный ответ. Рециркуляция лимфоцитов.
2. Происхождение ( образование ) клеток иммунной системы. Функции клеток иммунной системы. Лимфопоэз. Бурса Фабрициуса.
3. Т-лимфоциты. Характеристика Т-лимфоцитов. Типы молекул на поверхности Т-лимфоцитов.
4. Популяции Т-лимфоцитов. Субпопуляции Т-лимфоцитов. СD4 Т-лимфоциты. СD8 Т-лимфоциты.
5. Функции Т-лимфоцитов. Активированные Т-лимфоциты. Цитокины.
6. В-лимфоциты. Характеристика В-лимфоцитов. Клетки памяти.
7. Функции В-лимфоцитов. Типы молекул на поверхности В-лимфоцитов.
8. Антигенпредставляющие клетки. Структура и функции органов иммунной системы.
9. Костный мозг. Функции костного мозга. Миеломоноцитопоэз.
10. Тимус (вилочковая железа). Тимоциты. Функции тимуса ( вилочковой железы ). Этапы отбора тимоцитов.
Происхождение ( образование ) клеток иммунной системы. Функции клеток иммунной системы. Лимфопоэз. Бурса Фабрициуса.
Лимфоциты, как и другие клетки крови, ведут свое происхождение из единой стволовой гемопоэтической клетки. Лимфопоэз имеет черты сходства с общим гемопоэзом: зависит от контактов клеток-предшественниц со стро-мальными клетками и от влияния цитокинов. Вместе с тем лимфопоэзу присущи некоторые особенности: возникновение в процессе пролиферации клеток-предшественниц широкого разнообразия Т- и В-лимфоцитов с разными антигенраспознающими рецепторами; гибель путем апоптоза значительной части пролиферирующих клеток-предшественниц; избирательное накопление в результате пролиферации клеток — потомков одной клетки, первоначально активированной при контакте с соответствующим антигеном, «клона» клеток, все клетки которого несут одинаковые по специфичности антигенраспознающие рецепторы.
Лимфоциты — это единственные клетки организма, способные специфически распознавать собственные и чужеродные антигены и отвечать активацией на контакт с конкретным антигеном. При весьма сходной морфологии малые лимфоциты делятся на две популяции, имеющие различные функции и продуцирующие разные белки (табл. 8.1).
Одна из популяций получила название В-лимфоциты, от названия органа «бурса Фабрициуса», где было впервые обнаружено созревание этих клеток у птиц. У человека В-лимфоциты созревают в костном мозге. В-лимфоциты распознают антигены специфическими рецепторами иммуно-глобулиновой природы, которые по мере созревания В-лимфоцитов появляются на их мембранах. Взаимодействие антигена с такими рецепторами является сигналом активации В-лимфоцитов и их дифференцировки в плазматические клетки, продуцирующие и секретирующие специфические для данного антигена антитела — иммуноглобулины.
Таблица 8.1. Классификация лимфоцитов человека
Другая популяция получила название Т-лимфоциты в связи с дифференцировкой их предшественников в тимусе. Эта популяция включает разные субпопуляции, среди которых наиболее отчетливо различаются Т-хелперы (CD4+) и цитотоксические Т-лимфоциты (CD8+). В отличие от В-лимфоцитов, способных распознавать и связывать белковые, полисахаридные и липопротеидные растворимые антигены, Т-лимфоциты могут распознавать только короткие пептидные фрагменты белковых антигенов на поверхности вспомогательных антигенпредставляющих клеток (дендритных, макрофагов или В-лимфоцитов) в комплексах с собственными молекулами главного комплекса гистосовместимости. Связывание антигена с антиген-распознающим рецептором на клеточной мембране индуцирует активацию Т-лимфоцита, которая проявляется продукцией и секрецией цитокинов, усиливающих процессы пролиферации и дифференцировки самих Т-лимфоцитов, В-лимфоцитов и макрофагов.
Т-лимфоциты выполняют важнейшую функцию специфического распознавания и связывания антигена, что ведет к их активации, пролиферации и дифференцировке разных субпопуляций Т-лимфоцитов, участвующих далее во всех формах иммунного ответа.
Главной функцией В-лимфоцитов также является специфическое распознавание антигена, которое ведет к их активации, пролиферации и дифференцировке в плазматические клетки — продуценты специфических антител — иммуноглобулинов, т. е. к гуморальному иммунному ответу. Чаще всего В-лимфоциты для развития гуморального иммунного ответа нуждаются в помощи Т-лимфоцитов в виде продукции активирующих цитокинов.
Имеется еще один тип лимфоцитов — большие гранулярные лимфоциты, отличающиеся от меньших по величине Т- и В-лимфоцитов не только особенностями структуры, но и отсутствием антигенраспознающего рецептора. Эти клетки получили название «естественные киллеры»: они способны убивать инфицированные разными вирусами клетки-мишени или опухолевые клетки (см. табл. 8.1).
Все процессы транспортировки лимфоцитов из первичных во вторичные органы иммунной системы, из вторичных органов иммунной системы в ткани зависят от взаимодействия особого типа цитокинов — хемокинов с соответствующими рецепторами на поверхности клеток. Кроме хемокинов миграция лимфоцитов зависит от присутствия на их поверхности адгезионных молекул, которым соответствуют молекулы-лиганды на поверхности эндотелиальных клеток сосудов, на поверхности макрофагов и дендритных клеток, на элементах внеклеточного матрикса.
– Также рекомендуем “Т-лимфоциты. Характеристика Т-лимфоцитов. Типы молекул на поверхности Т-лимфоцитов..”
Источник
Характеристика иммунокомпетентных клеток
Основными клетками, осуществляющими иммунные реакции, являются Т- и В-лимфоциты (и производные последних – плазмоциты), макрофаги, а также ряд взаимодействующих с ними клеток (тучные клетки, эозинофилы и др.).
Лимфоциты
Популяция лимфоцитов функционально неоднородна. Различают три основных вида лимфоцитов: Т-лимфоциты, В-лимфоциты и так называемые нулевые лимфоциты (0-клетки). Лимфоциты развиваются из недифференцированных лимфоидных костномозговых предшественников и при дифференцировке получают функциональные и морфологические признаки (наличие маркеров, поверхностных рецепторов), выявляемые иммунологическими методами. 0-лимфоциты (нулевые) лишены поверхностных маркеров и рассматриваются как резервная популяция недифференцированных лимфоцитов.
Т-лимфоциты — самая многочисленная популяция лимфоцитов, составляющая 70—90% лимфоцитов крови. Они дифференцируются в вилочковой железе — тимусе (отсюда их название), поступают в кровь и лимфу и заселяют Т-зоны в периферических органах иммунной системы — лимфатических узлах (глубокая часть коркового вещества), селезенке (периартериальные влагалища лимфоидных узелков), в одиночных и множественных фолликулах различных органов, в которых под влиянием антигенов образуются Т-иммуноциты (эффекторные) и Т-клетки памяти.
Для Т-лимфоцитов характерно наличие на плазмолемме особых рецепторов, способных специфически распознавать и связывать антигены. Эти рецепторы являются продуктами генов иммунного ответа [Ir-гены, (immune response)]. Т-лимфоциты обеспечивают клеточный иммунитет, участвуют в регуляции гуморального иммунитета, осуществляют продукцию цитокинов при действии антигенов.
В популяции Т-лимфоцитов различают несколько функциональных групп клеток: цитотоксические лимфоциты (Тц), или Т-киллеры (Тк), Т-хелперы (Тх), Т-супрессоры (Тс). Тк участвуют в реакциях клеточного иммунитета, обеспечивая разрушение (лизис) чужеродных клеток и собственных измененных клеток (например, опухолевых клеток). Рецепторы позволяют им распознавать белки вирусов и опухолевых клеток на их поверхности. При этом активизация Тц (киллеров) происходит под влиянием антигенов гистосовместимости на поверхности чужеродных клеток.
Кроме того, Т-лимфоциты участвуют в регуляции гуморального иммунитета с помощью Тх и Тс. Тх стимулируют дифференцировку В-лимфоцитов, образование из них плазмоцитов и продукцию иммуноглобулинов (Ig). Tx имеют поверхностные рецепторы, которые связываются с белками на плазмолемме В-клеток и макрофагов, стимулируя Тх и макрофаги к пролиферации, продукции интерлейкинов (пептидных гормонов), а В-клетки — к продукции антител.
Таким образом, главной функцией Тх является распознавание чужеродных антигенов (представляемых макрофагами), секреция интерлейкинов, стимулирующих В-лимфоциты и другие клетки для участия в иммунных реакциях.
Снижение в крови числа Тх ведет к ослаблению защитных реакций организма (эти лица более подвержены инфекциям). Отмечено резкое снижение числа Тх у лиц, инфицированных вирусом СПИДа.
Тс способны ингибировать активность Тх, В-лимфоцитов и плазмоцитов. Они участвуют в аллергических реакциях, реакциях гиперчувствительности. Тс подавляют дифференцировку В-лимфоцитов.
Одной из основных функций Т-лимфоцитов является продукция цитокинов, которые оказывают стимулирующее или тормозящее влияние на клетки, участвующие в иммунном ответе (хемотаксические факторы, макрофаги ингибирующий фактор — МИФ, неспецифические цитотоксические вещества и др.).
Натуральные киллеры. Среди лимфоцитов в крови, кроме вышеописанных Тц, выполняющих функцию киллеров, имеются так называемые натуральные киллеры (Нк, NK), которые также участвуют в клеточном иммунитете. Они образуют первую линию защиты против чужеродных клеток, действуют немедленно, быстро разрушая клетки. Нк в собственном организме разрушают опухолевые клетки и клетки, инфицированные вирусом. Тц образуют вторую линию защиты, так как для их развития из неактивных Т-лимфоцитов требуется время, поэтому они вступают в действие позже Нк. Нк — это большие лимфоциты диаметром 12—15 мкм, имеют дольчатое ядро и азурофильные гранулы (лизосомы) в цитоплазме.
В-лимфоциты являются основными клетками, участвующими в гуморальном иммунитете. У человека они образуются из СКК красного костного мозга, затем поступают в кровь и далее заселяют В-зоны периферических лимфоидных органов — селезенки, лимфатических узлов, лимфоидные фолликулы многих внутренних органов. В крови их содержится 10—30% от всей популяции лимфоцитов.
Для В-лимфоцитов характерно наличие на плазмолемме поверхностных иммуноглобулиновых рецепторов (SIg или MIg) для антигенов. Каждая В-клетка содержит 50000…150000 антигенспецифических молекул SIg. В популяции В-лимфоцитов находятся клетки с различными SIg: большинство (⅔) содержат IgM, меньшее число (⅓) — IgG и около 1—5 % — IgA, IgD, IgE. В плазмолемме В-лимфоцитов имеются также рецепторы для комплемента (С3) и Fc-рецепторы.
При действии антигена В-лимфоциты в периферических лимфоидных органах активизируются, пролиферируют, дифференцируются в плазмоциты, активно синтезирующие антитела различных классов, которые поступают в кровь, лимфу и тканевую жидкость.
Развитие Т- и В-лимфоцитов
Родоначальником всех клеток иммунной системы является кроветворная стволовая клетка (СКК). СКК локализуются в эмбриональном периоде в желточном мешке, печени, селезенке. В более поздний период эмбриогенеза они появляются в костном мозге и продолжают пролиферировать в постнатальной жизни. Из СКК в костном мозге образуется клетка—предшественник лимфопоэза (лимфоидная мультипотентная родоначальная клетка), которая генерирует два типа клеток: пре-Т-клетки (предшественники Т-клеток) и пре-В-клетки (предшественники В-клеток).
Дифференцировка Т-лимфоцитов
Пре-Т-клетки мигрируют из костного мозга через кровь в центральный орган иммунной системы — вилочковую железу (тимус). Еще в период эмбрионального развития в вилочковой железе создается микроокружение, имеющее значение для дифференцировки Т-лимфоцитов. В формировании микроокружения особая роль отводится ретикулоэпителиальным клеткам этой железы, способным к продукции ряда биологически активных веществ. Мигрирующие в вилочковую железу пре-Т-клетки приобретают способность реагировать на стимулы микроокружения. Пре-Т-клетки в вилочковой железе пролиферируют, трансформируются в Т-лимфоциты, несущие характерные мембранные антигены (CD4+, CD8+). Т-лимфоциты генерируют и «поставляют» в кровообращение и в тимусзависимые зоны периферических лимфоидных органов 3 типа лимфоцитов: Тц, Тх и Тс. Мигрирующие из вилочковой железы «девственные» Т-лимфоциты (виргильные Т-лимфоциты) являются короткоживущими. Специфическое взаимодействие с антигеном в периферических лимфоидных органах служит началом процессов их пролиферации и дифференцировки в зрелые и долгоживущие клетки (Т-эффекторные и Т-клетки памяти), составляющие большую часть рециркулирующих Т-лимфоцитов.
Из вилочковой железы мигрируют не все клетки. Часть Т-лимфоцитов погибает. Существует мнение, что причиной их гибели служит присоединение антигена к антигенспецифическому рецептору. В вилочковой железе нет чужеродных антигенов, поэтому данный механизм может служить для удаления Т-лимфоцитов, способных реагировать с собственными структурами организма, т.е. выполнять функцию защиты от аутоиммунных реакций. Гибель части лимфоцитов является генетически запрограммированной (апоптоз).
Дифференцировочные антигены Т-клеток. В процессе дифференцировки лимфоцитов на их поверхности появляются специфические мембранные молекулы гликопротеидов. Такие молекулы (антигены) можно обнаружить с помощью специфических моноклональных антител. Получены моноклональные антитела, которые реагируют лишь с одним антигеном клеточной мембраны. С помощью набора моноклональных антител можно идентифицировать субпопуляции лимфоцитов. Имеются наборы антител к дифференцировочным антигенам лимфоцитов человека. Антитела составляют относительно немного групп (или «кластеров»), каждая из которых узнает один единственный белок клеточной поверхности. Создана номенклатура дифференцировочных антигенов лейкоцитов человека, выявляемых моноклональными антителами. Эта CD-номенклатура (CD — cluster of differentiation — кластер дифференцировки) базируется на группах моноклональных антител, реагирующих с одними и теми же дифференцировочными антигенами.
Получены многоклональные антитела к ряду дифференцировочных антигенов Т-лимфоцитов человека. При определении общей популяции Т-клеток могут быть использованы моноклональные антитела специфичностей CD (CD2, CD3, CDS, CD6, CD7).
Известны дифференцировочные антигены Т-клеток, которые характерны либо для определенных стадий онтогенеза, либо для различающихся по функциональной активности субпопуляций. Так, CD1 — маркер ранней фазы созревания Т-клеток в вилочковой железе. В процессе дифференцировки тимоцитов на их поверхности экспрессируются одновременно маркеры CD4 и CD8. Однако в последующем маркер CD4 исчезает с части клеток и сохраняется только на субпопуляции, переставшей экспрессировать антиген CD8. Зрелые CD4+ клетки являются Тх. Антиген CD8 экспрессируется примерно на ⅓ периферических Т-клеток, которые созревают из CD4+/CD8+ Т-лимфоцитов. Субпопуляция CD8+ Т-клеток включает цитотоксические и супрессорные Т-лимфоциты. Антитела к гликопротеинам CD4 и CD8 широко используются для того, чтобы различать и разделять Т-клетки соответственно на Тх и Тц.
Кроме дифференцировочных антигенов, известны специфические маркеры Т-лимфоцитов.
Т-клеточные рецепторы для антигенов представляют собой антителоподобные гетеродимеры, состоящие из полипептидных α- и β-цепей. Каждая из цепей имеет длину в 280 аминокислот, большая внеклеточная часть каждой цепи свернута в два Ig-подобных домена: один вариабельный (V) и один константный (С). Антителоподобный гетеродимер кодируется генами, которые собираются из нескольких генных сегментов в процессе развития Т-клеток в вилочковой железе.
Дифференцировка В-лимфоцитов
Предшественники В-клеток (пре-В-клетки) развиваются в дальнейшем у птиц в фабрициевой сумке (bursa), откуда произошло название В-лимфоциты, у человека и млекопитающих — в костном мозге.
Сумка Фабрициуса (bursa Fabricii) — центральный орган иммунопоэза у птиц, где происходит развитие В-лимфоцитов, находится в области клоаки. Для ее микроскопического строения характерно наличие многочисленных складок, покрытых эпителием, в которых расположены лимфоидные узелки, ограниченные мембраной. В узелках содержатся эпителиоциты и лимфоциты на различных стадиях дифференцировки. В период эмбриогенеза в центре фолликула формируется мозговая зона, а на периферии (снаружи от мембраны) — корковая зона, в которую, вероятно, мигрируют лимфоциты из мозговой зоны. В связи с тем что в сумке Фабрициуса у птиц образуются исключительно В-лимфоциты, она является удобным объектом для изучения строения и иммунологических характеристик этого вида лимфоцитов. Для ультрамикроскопического строения В-лимфоцитов характерно наличие в цитоплазме групп рибосом в виде розеток. Эти клетки имеют более крупные ядра и менее плотный хроматин, чем у Т-лимфоцитов, в связи с увеличением содержания эухроматина.
В-лимфоциты отличаются от других типов клеток способностью синтезировать иммуноглобулины. Зрелые В-лимфоциты экспрессируют Ig на клеточной мембране. Такие мембранные иммуноглобулины (MIg) функционируют как антигенспецифические рецепторы.
Пре-В-клетки синтезируют внутриклеточный цитоплазматический IgM, но не имеют поверхностных иммуноглобулиновых рецепторов. Костномозговые виргильные В-лимфоциты имеют IgM-рецепторы на своей поверхности. Зрелые В-лимфоциты несут на своей поверхности иммуноглобулиновые рецепторы различных классов — IgM, IgG и др.
Дифференцированные В-лимфоциты поступают в периферические лимфоидные органы, где при действии антигенов происходят пролиферация и дальнейшая специализация В-лимфоцитов с образованием плазмоцитов и В-клеток памяти (ВП).
В ходе своего развития многие В-клетки переключаются с выработки антител одного класса на выработку антител других классов. Этот процесс называется переключением класса. Все В-клетки начинают свою деятельность по синтезу антител с выработки молекул IgM, которые встраиваются в плазматическую мембрану и служат рецепторами для антигена. Затем, еще до взаимодействия с антигеном, большая часть В-клеток переходит к одновременному синтезу молекул IgM и IgD. Когда виргильная В-клетка переходит от выработки одного лишь мембраносвязанного IgM к одновременному синтезу мембраносвязанных IgM и IgD, переключение происходит, вероятно, благодаря изменению процессинга РНК.
При стимуляции антигеном некоторые из этих клеток активируются и начинают выделять антитела IgM, преобладающие в первичном гуморальном ответе.
Другие стимулированные антигеном клетки переключаются на выработку антител классов IgG, IgE или IgA; В-клетки памяти несут эти антитела на своей поверхности, а активные В-клетки их секретируют. Молекулы IgG, IgE и IgA в совокупности называются антителами вторичных классов, так как они, по-видимому, образуются только после антигенной стимуляции и преобладают во вторичных гуморальных ответах.
При помощи моноклональных антител удалось выявить определенные дифференцировочные антигены, которые еще до появления цитоплазматических µ-цепей позволяют отнести несущий их лимфоцит к В-клеточной линии. Так, антиген CD19 является самым ранним маркером, позволяющим отнести лимфоцит к В-клеточному ряду. Он присутствует на пре-В-клетках в костном мозге, на всех периферических В-клетках.
Антиген, выявляемый моноклональными антителами группы CD20, специфичен для В-лимфоцитов и характеризует более поздние стадии дифференцировки.
На гистологических срезах антиген CD20 выявляется на В-клетках герминативных центров лимфоидных узелков, в корковом веществе лимфатических узлов. В-лимфоциты несут также ряд других (например, CD24, CD37) маркеров.
Различают антигеннезависимую и антигензависимую дифференцировку и специализацию В- и Т-лимфоцитов.
Антигеннезависимая пролиферация и дифференцировка генетически запрограммированы на образование клеток, способных давать специфический тип иммунного ответа при встрече с конкретным антигеном благодаря появлению на плазмолемме лимфоцитов особых «рецепторов». Она совершается в центральных органах иммунитета (тимус, костный мозг или фабрициева сумка у птиц) под влиянием специфических факторов, вырабатываемых клетками, формирующими микроокружение (ретикулярная строма или ретикулоэпителиальные клетки в тимусе).
Антигензависимая пролиферация и дифференцировка Т- и В-лимфоцитов происходят при встрече с антигенами в периферических лимфоидных органах, при этом образуются эффекторные клетки и клетки памяти (сохраняющие информацию о действовавшем антигене).
Образующиеся Т-лимфоциты составляют пул долгоживущих, рециркулирующих лимфоцитов, а В-лимфоциты — короткоживущих клеток.
Некоторые термины из практической медицины:
- вакцина (vaccinum; лат. vaccinus коровий) — препарат, получаемый из живых аттенуированных штаммов или убитых культур микроорганизмов, их токсинов или антигенов, предназначенный для активной иммунизации людей и животных;
- анавакцина (anavaccinum; ана- + вакцина) — убитая вакцина, приготовленная путем продолжительного воздействия на взвесь микробов раствора формалина;
- вакцинация (vaccinatio; франц. vaccination) — метод создания активного иммунитета против инфекционной болезни путем введения вакцины в организм человека или животного;
- иммунизация (1) (immunisatio; лат. immunis свободный, избавленный от чего-либо; син.: прививки) — противоэпидемическое мероприятие, основанное на регламентированном по времени и возрастным группам применении средств специфической профилактики инфекционных болезней;
- иммунизация (2) (immunisatio) введение вакцин (анатоксинов) животным-продуцентам и людям – донорам крови с целью получения от них иммунных сывороток (иммуноглобулинов);
- иммунитет активный иммунитет, возникающий в результате иммунного ответа организма на введение антигена;
- иммунитет пассивный иммунитет, возникающий при передаче антител в форме антисыворотки или иммуноглобулинов от иммунизированного донора, а также от матери через плаценту или с молозивом;
- иммунологическая недостаточность (син.: иммунодефицитное состояние, иммунопарез, иммунодефицит) — снижение или отсутствие иммунного ответа вследствие дефекта системы иммунитета;
Источник