Микроокружение для дифференцирующих лимфоцитов в тимусе

?Эпителиоретикулярные клетки Тимуса классифицируются на:
+секреторные
+клетки няньки
=сенсорные клетки
+периваскулярные
=интерстициальные
?В тимусе происходит:
=антиген-зависимая пролиферация и дифференцировка Т- и В-лимфоцитов
=антиген-независимая пролиферация и дифференцировка В-лимфоцитов
+антиген-независимая пролиферация и дифференцировка Т- лимфоцитов
=антиген-зависимая пролиферация естественных киллеров
=антиген-зависимая пролиферация и дифференцировка макрофагов
?При клеточном иммунитете эффекторными клетками являются:
=В-лимфоциты
+Т-лимфоциты цитотоксические
=Т-лимфоциты супрессоры
=Т-лимфоциты хелперы
=лимфоциты
?Для лимофузла характерны следующие образования:
=лимфатические узелки с центральной артерией
+лимфатические узелки, мозговые тяжи, синусы
=корковое и мозговое вещество без лимфатических узелков
=лимфатические узелки и многослойный неороговевающий эпителий
=лимфатические узелки, однослойный эпителий и крипты
?В селезенке происходит:
+антиген-зависимая пролиферация и дифференцировка Т- и В-лимфоцитов
=антиген-независимая пролиферация и дифференцировка В-лимфоцитов
=антиген-независимая пролиферация и дифференцировка Т- лимфоцитов
=антиген-зависимая пролиферация естественных киллеров
=антиген-зависимая пролиферация и дифференцировка макрофагов
?Красный костный мозг состоит из:
+гемопоэтического компонента
+стромального компонента
=интестициального компонента
+сосудистого компонента
=секреторного компонента
?К периферическим органам иммуногенеза относятся:
+селезенка
+лимфоузлы
+лимфатические узелки пищеварительного тракта и дыхательных путей
=тимус
?Для селезенки характерны следующие образования:
+лимфатические узелки с центральной артерией
=лимфатические узелки, мозговые тяжи, синусы
=корковое и мозговое вещество без лимфатических узелков
=лимфатические узелки и многослойный неороговевающий эпителий
=лимфатические узелки, однослойный эпителий и крипты
?Функциями селезенки являются:
+кроветворение
+уничтожение старых и поврежденных эритроцитов и тромбоцитов
+защитная реакция на проникающие в кровь антигены
=уничтожение старых и поврежденных нейтрофилов
?В красном костном мозге происходит:
=антиген-зависимая пролиферация и дифференцировка Т- и В-лимфоцитов
+антиген-независимая пролиферация и дифференцировка В-лимфоцитов
=антиген-независимая пролиферация и дифференцировка Т- лимфоцитов
=антиген-зависимая пролиферация естественных киллеров
=антиген-зависимая пролиферация и дифференцировка макрофагов
?Центральными органами иммуногенеза являются:
+тимус
=лимфоузлы
+красный костный мозг
=селезенка
=миндалины
?Для тимуса характерны следующие образования:
=лимфатические узелки с центральной артерией
=лимфатические узелки, мозговые тяжи, синусы
+корковое и мозговое вещество без лимфатических узелков
=лимфатические узелки и многослойный неороговевающий эпителий
?Клетками микроокружения для лимфоцитов в тимусе являются:
+эпителиоретикулоциты
=фибробласты
=макрофаги
=гранулоциты
?В состав ацинуса поджелудочной железы входят:
+экзокринные панкреоциты
=инсулоциты
+клетки вставочного отдела (центроацинозные клетки)
=миоэпителиальные клетки
?Периферическими органами иммуногенеза являются:
+лимфоузлы
+селезенка
+миндалины
=тимус
?В реакциях клеточного иммунитета участвуют:
=макрофаги
=В лимфоциты
+Т киллеры
=плазматические клетки
?В реакциях специфического клеточного иммунитета участвуют:
=В клетки
+Т-киллеры
+Т-хелперы
=плазмоциты
?Антиген-зависимая дифференцировка Т- и В-лимфоцитов происходит в:
=красном костном мозге
=тимусе
+селезенке
+лимфоузлах
?Клетки, синтезирующие антитела, образуются из:
=В-лимфоцитов памяти;
=Т-лимфоцитов;
+В-лимфоцитов;
=макрофагов.
?Антиген-независимая дифференцировка В-лимфоцитов происходит в:
+красном костном мозге
=тимусе
=печени
=периферических лимфоидных органов
?Антиген-зависимая дифференцировка Т- и В-лимфоцитов происходит в:
=красном костном мозге
=тимусе
=печени
+периферических лимфоидных органах
?Т-киллеры осуществляют:
=синтез антител
+лизис клеток с антигенной информацией
=фагоцитоз клеток с антигенной информацией
=выделение медиаторов, привлекающих макрофаги и гранулоциты
?Клетками микроокружения для гемопоэтических клеток в красном костном мозге являются:
+ретикулярные
=остеогенные
=макрофаги
=адипоциты
?Гематотимусный барьер не включает:
=цитоплазму эндотелиоцита с его базальной мембраной
=периваскулярное пространство
=цитоплазму эпителиоретикулоцита
=базальную мембрану эпителиоретикулоцита
+цитоплазму тимоцитов
?Функции селезенки:
+участие в формировании гуморального и клеточного иммунитета
+депонирование крови
+накопление тромбоцитов
+разрушение старых и поврежденных эритроцитов и тромбоцитов
=антигеннезависимая рекция Т-лимфоцитов
?Лимфатические синусы бывают:
+мозговой
=корковый
+субкапсулярный
+промежуточный
=паракортикальный
?Основная функция лимфы:
=трофическая
=дыхательная
+иммунная
=секреторная
=дыхательная
КОНТРОЛЬНЫЙ ТЕСТ
Источник
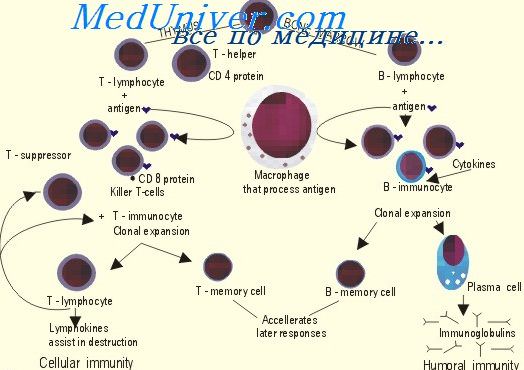
Образование Т-клеток в тимусе. Заселение тимуса предшественниками лимфоцитовОбразование Т-клеток в тимусе происходит из потомков костномозговых стволовых кроветворных клеток. Эмигрируя из костного мозга, они попадают по току крови в тимус и проникают в его ткань в области коркового слоя. В тимусе они активно пролиферируют и подвергаются дифференцировке и коммитированию. Их потомки перемещаются в корковый слой и либо гибнут внутри тимуса, либо покидают его, вновь поступая в кровоток. Стадии дифферепцировки, которые проделывают предшественники Т-клеток в тимусе, имеют антигенные, функциональные и морфологические маркеры. Наименее зрелые предшественники характеризуются высокой чувствительностью к кортизону, высокой концентрацией антигенов TL и Ly и низкой концентрацией Н2-антигенов. Морфологически эти клетки являются малыми лимфоцитами, содержащими монорибосомы и относительно мало органоидов. Они дают начало пролиферирующим в тимусе большим лимфоцитам с резко увеличенным количеством рибосом, собранных в полисомы в виде розеток. Наиболее зрелые в пределах тимуса Т-клетки находятся в его мозговом слое. Они резистентны к кортизону, имеют малые концентрации TL- и 8-антигенов, но много Н2-антигена. Морфологически — это малые лимфоциты, но содержащие полирибосомы и волокнистые цитоплазматические структуры. Таких клеток в тимусе содержится около 5% от всех тимоцитов, и именно они могут выходить из тимуса в кровоток и осуществлять колонизацию периферических лимфоидных органов в качестве антигенраспознающих, уже коммитированных Т-клеток. Окончательное дозревание Т-клеток происходит только после их попадания, во вторичные лимфоидные органы.
Гибель подавляющей части тимоцитов внутри тимуса позволяет предполагать, что избыточная пролиферация в нем Т-предшественников связана с апробированием их свойств и отбором еще до встречи с антигенами; последние практически не попадают в тимус благодаря наличию специального барьера. При заселении тимуса предшественниками Т-клеток ориентирами для них служат не лимфоциты (тимоциты) и не макрофаги (которые в тимусе крайне малочисленны), а клетки его стромы. На это указывают результаты типирования клеток в гетеротопных трансплантатах тимуса (Дидух, Фриденштейн, 1970). Стромальные клетки тимуса — ретикулярные клетки, или клетки его эктодермального эпителия,— создают и то микроокружение, которое необходимо для дифференцировки Т-клеток: в отсутствие тимической стромы Т-клетки в организме не образуются. Однако результаты, полученные на радиохимергх и при гетеротопной трансплантации тимуса, показали, что свойства образующихся в тимусе Т-клеток определяются генетической структурой заселяющих его предшественников, а не стромальными клетками, образующими территорию, на которой происходит их дифференцировка. Действительно, в случаях, когда тимэктомированный реципиент и донор тимуса, используемого для гетеротопной трансплантации, различаются по антигенам, тимоциты, которые образуются в пересаженном тимусе, имеют 6-антиген реципиента, а в случае, когда по 6-антигенам различаются донор клеток костного мозга и облученный реципиент, тимоциты радиохимеры имеют 6-антиген донора (Metcalf, Moore, 1971). Это показывает, что роль тимического микроокружения как индуктора образования Т-клеток состоит в создании возможности для реализации тех генетически закодированных в потомках стволовых кроветворных клеток свойств, которые характеризуют дифференцировку Т-клеток. Важный фактор тимического микроокружения — это секретируемый его стромальными клетками полипептид — тимозин (м. в. 35 000). Тимозин индуцирует созревание Т-клеток из костномозговых предшественников (в частности, синтез 6-антигена) как in vivo, так и in vitro (Scheid e. a., 1973). Исчерпывается ли специфическое действие тимического микроокружения секрецией тимозина, пока не выяснено. – Также рекомендуем “Колонизация периферических лимфоидных органов. Репопуляция лимфоцитов” Оглавление темы “Регуляция формирования антител”: |
Источник
- Иммунология и аллергология
- Иммунология
- Иммунология: Учебник
Два наблюдения: необходимость тимуса для накопления функционально зрелых Т-клеток на периферии и высокая гибель активно пролиферирующих тимоцитов in situ, указывают на тот факт, что в тимусе происходят те основные события, которые определяют полноценность работы всей Т-системы. Подобная феноменология требовала выяснения, по крайней мере, двух вопросов: каков биологический смысл массовой гибели клеток в тимусе и как формируются субпопуляции клоноспецифических Т- клеток (Т- киллеров, Т-хелперов) в органе?
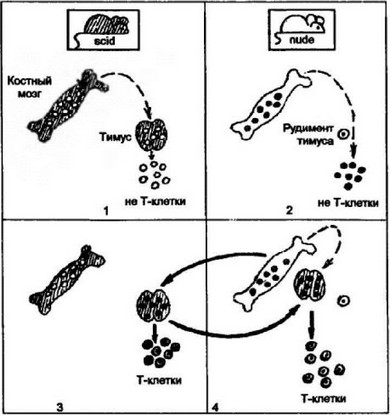
Рис. 7.1. Значение эпителиального микроокружения тимуса для дифференцирован костномозговых предшественников до зрелых Т-клеток.
Мутантные мыши scid имеют полноценный тимус, однако Т-клетки не образуются, так как имеется дефект рекомбинации генов для антиген распознающих рецепторов (1). Другая мутантная линия мышей — nude — также неспособна к формированию Т-клеток из-за отсутствия тимуса: у этой линии мышей обнаруживается только рудимент органа (2). Трансплантация клеток костного мозга мышей nude мышам scid обеспечивает развитие Т-клеток (3). Опыт демонстрирует наличие у мышей nude полноценных костномозговых предшественников Т-клеток. При трансплантации мышам nude тимуса от мышей scid восстанавливается образование Т-клеток у nude-хозяина. Эта форма трансплантации показывает роль эпителиального микроокружения тимуса для нормального развития костномозговых предшественников до зрелых Т-клеточных форм
Этапы внутритимусной дифферен цировки клеток от мигрировавшего в орган костномозгового предшественника (пре-Т-клет- ки) до зрелого Т-лимфоцита, покидающего тамус, связаны с изменением экспрессии фенотапических, Т-клеточных маркеров. Основными из них являются: CD4 — корецептор Т-хелперов, CD8 — корецептор цитотоксических Т-лимфоцитов (ЦТЛ; Т-киллеров) и сфТКР — Т-клеточный антигенраспознающий рецептор. Специфическая комбинация этих поверхностных молекул может быть использована в качестве маркеров дифферен цировки клеток в тимусе (табл. 7.1).
Первые мигранты из костного мозга — пре-Т-клетки — представляют собой лимфобласты, имеющие определенный набор
Таблица 7.1
Фенотипические маркеры дифференцирующихся тимоцитов
| Этап | Маркеры | Тип дифференцирующихся тимоцитов | |
| корсцепторы Т-клеток | Т-клеточный рецептор | ||
| I | CD4 CD8- | apTKP- | двойной негатив |
| 2 | cm* CDs* | apTKP” | |
| 3 | CD4+ CD8+ | aflTKP* | двойной позитив |
| 4 | CD4+ CD8+ | apTKP+ | двойной ПОЗИТИВ |
| 5 | CD4+ CDS’ | apTKP+ | одинарный позитив |
| CD4- CD8+ | apTKP* | одинарный позитив | |
поверхностных молекул, но лишенных основных маркеров диф- ференцировки: CD4 и CD8 (рис. 1.2). Отсюда и их название — “двойные негативы11. Они заселяют верхнюю часть коры тимуса, расположенные, непосредственно под капсулой органа — субкапсулярную область (рис. 7.3). В полностью развитом тимусе двойные негативные клетки составляют незначительный пул (всего около 5% от общего числа тимоцитот?
Взаимодействие раннего предшественника со стромой субкапсулярной области приводит к экспрессии первого специфического маркера Т-клеток: CD2 — у людей и Thy-1 — у мышей. Обе молекулы обладают адгезивными свойствами и относятся к суперсемейству иммуноглобулинов. Thy-1 является исключительным маркером Т-клеток мышей и сохраняется на всех стадиях диф- ференцировки данных клеток, хотя уровень его экспрессии у более зрелых тимоцитов и периферических Т-клеток снижен по сравнению с ранними предшественниками.
Тимоцитарные бласты субкапсулярной области, находясь в тесном контакте с эпителиальными клетками-кормилицами, ак-
Рис. 7.2. ЭпшянттшеэаикпсойдяФфереяцщхяпТ-клстмс.
На первом этапе из плюри потентной стволовой кроветворной клетки (СКК) костного мозга, обеспечивающей миело- и лимфопоэз, образуется общий для Т- н В-лимфоцитов предшественник — ПрЛЦ. Ближайший потомок ПрЛЦ — проти- моцит, или коммитированный предшественник Т-клеток (пре-Т-кл.). Характерным маркером пре-Т-клеток костного мозга является один из антигенов мозга (AM). Первые мигрирующие в субкапсулярную область тимуса пре-Т-клетки теряют AM, но приобретают типичный маркер тимоцитов и периферических Т-клеток — Thy-1. Тимоциты субкапсулярной зоны являются в основном двойными негативами и не экспрессируют Т-клеточный рецептор (ТКР). Фенотип таких клеток — CD4″CD8~a0TKP Постепенно, по мере перемещения в корковый слой тимоциты начинают экспрессию как CD4-, так и С08-корецепторов, а также сфТКР. Медуллярная зона тимуса — место локализации властных форм с фенотипом, характерным для самостоятельных субпопуляций Т-клеток (CD4+CD8’a0TKP+ — Т-хелпе- ры/индукторы; CD4’CD8+apTKP*). Формированием различных субпопуляций завершается внутритимусная дифференцировка тимоцитов. Образовавшиеся субпопуляции мигрируют в периферические лимфоидные органы (лимфатические узлы, селезенку и др.)
Рис. 7.3. Локализация дифференцирующихся тнмоцнтоь и различных областях пща.
Пре-Т- клетки, мигрирующие из костного мозга в тимус, заселяют субкапсулярную зону тимуса. Клетки этой стадии развития представляют собой незрелые формы, которые не экспрессируют ТКР и корецепторы (фенотип CD4CD8TKP). В результате их взаимодействия с эпителиальными клетками-кормилицами, представленными в субкапсуле, начинается процесс внутритимусной дифференциров- ки. В промежуточной зоне между субкапсулой и корой, а также частично в коре, появляются тимоциты с умеренной экспрессией корецепторов и (3-цепи ТКР (фенотип CD4±CD8±pTKP±). Под влиянием эпителиальных клеток коры проходит дальнейшее созревание тимоцитов, что регистрируется по усилению экспрессии корецепторов CD4 и CD8 и полноценой экспрессии ТКР (фенопип СШ’ХОв^офТКР*). Последующая дифференцмровка осуществляется в переходной зоне кортико-медуллярного соединения под влиянием дендритных клеток и макрофагов. В результате образуются сублопуляции Т-киллеров/супрессоров (фенотип CD4′ CD8+apTKP+) и Т-хелперов/индукторов (фенотип CD4+CD8’apTKP+)
тивно пролиферируют и завершают свой путь развития в данной области умеренной экспрессией CD4 и CD8. Тимоциты с фенотипом CD4*, CDS*, рТКР* перемещаются в корковый слой.
Клетки коры — это в основном малые, плотно упакованные тимоциты. Они находятся в непосредственных контактных отношениях с кортикальными эпителиальными клетками, обладающими разветвленными выростами, которые окружают тимоциты. Выраженная экспрессия на поверхности эпителиальных клеток молекул 1 и II классов МНС является определяющим фактором положительной селекции клеток тимуса (см. ниже). В результате взаимодействия эмигрантов из субкапсулярной области с эпителиальными клетками коры происходит активная, одновременная экспрессия на поверхности тимоцитов двух корецепторов — CD4 и CD8, что дало названием кортикальным клеткам “двойные позитивы”. При этом двойные позитивные тимоциты становятся обладателями полноценных арТКР. Таким образом основной фенотип кортикальных тимоцитов — CD4+, CD8+, арТКР4′.
Факт прохождения положительной селекции в коре определяет дальнейшую дифференцировку на субпопуляпии CD4+CD8′ и CD4″CD8+, которая осуществляется в основном в переходной области — кортико-медуллярном соединении под влиянием взаимодействия с дендритными клетками и макрофагами. Эти клетки получили название “одинарных позитивов”. Здесь же проходит и отрицательная селекция тимоцитов (см.ниже). В результате прошедших дифференцировочных событий в медуллярной области накапливаются клетки с фенотипом CD4+CD8’TKP+ (Т-хелгтеры) и CD4″CD8*TKP+ (Т-киллеры), часть которых мигрирует в периферические лимфоидные органы.
Источник: В.Г. Галактионов., «Иммунология: Учебник» 1998
А так же в разделе «Этапы внутритимусной дифференцирован лимфоцитов »
- Основная феноменология
- 7.1.3. Реорганизация генов Т-клеточного рецептора в процессе дифференцнровки тимоцитов
- Положительная селекция клеток в тимусе
- Отрицательная cejieKuun клеток в тимусе
- 7.1.4.4. Возможные механизмы положительной и отрицательной селекции клеток в тимусе
Источник