Лимфоциты в селезенке локализованы

Т- и В-лимфоциты, которые покидают соответственно вилочковую железу и костный мозг, находятся на ранней стадии иммунологического созревания. Когда антигенпрезентирующие клетки представляют антиген этим Т- и В-клеткам в первый раз, например, в лимфатическом узле или селезенке, лимфоциты трансформируются в Т- или В-иммунобласты.
Иммунобласты являются самыми крупными лимфоидными клетками. Они имеют светлое ядро с тонкоструктурированным хроматином и крупными одиночными ядрышками. Цитоплазма иммунобластов выглядит как широкий базофильный ободок. Т-иммунобласты либо выполняют свою эффекторную Т-клеточную функцию и погибают, либо становятся Т-клетками памяти. Клетки последнего типа более сильно и интенсивно отвечают на стимуляцию при контакте с тем же самым антигеном.
В-иммунобласты превращаются в плазматические клетки, проходя перед этим стадию плазмоцитоидных клеток. Первоначально плазмоцитоидные клетки имеют морфологические признаки, промежуточные между малыми лимфоцитами и зрелыми плазматическими клетками, на этой стадии они секретируют преимущественно IgM. При развитии размер клеток увеличивается, и они приобретают черты типичных плазматических клеток, продуцирующих иммуноглобулины различных классов.
Плазматические клетки крупнее, чем лимфоциты. Они имеют эксцентрично расположенное круглое ядро. Цитоплазма плазматических клеток строго базофильна, за исключением светлой перинуклеарной зоны, занятой комплексом Гольджи. При первичном иммунном ответе плазмоцито-идные лимфоциты преобладают, а ассоциированная с ними продукция иммуноглобулинов невелика.
Пути созревания В-лимфоцитов в лимфатическом узле после выхода из костного мозга
В лимфатических узлах и других лимфоидных органах, прежде всего селезенке, в ответ на антигенную стимуляцию Т-зависимыми антигенами образуются зародышевые центры. Первоначально В-клетки, из которых формируется зародышевый центр, получают активирующий сигнал вне фолликулов, в Т-клеточных зонах от интердигитальных дендритных клеток и Т-хелперов. Каждый фолликул колонизируют в среднем три В-клеточных бласта. Эти бласты подвергаются массивной клональной экспансии, и в них активизируется механизм соматических гипермутаций, который действует на вариабельные области генов иммуноглобулинов.
Зрелые зародышевые центры разделены на темную и светлую зоны. Темную зону занимают пролиферирующие бласты, называемые центробластами, затем из них формируются центроциты, которые не находятся в митотическом цикле и вытесняются в светлую зону. Два типа этих лимфоидных клеток известны как клетки фолликулярных центров. Центробласты — крупные клетки, но обычно мельче иммунобластов.
Светлая зона содержит сеть фолликулярных дендритных клеток, которые имеют способность поглощать и процессировать (обрабатывать) антиген. Антигенные детерминанты могут удерживаться на поверхности дендритных клеток более года или в форме иммунного комплекса, или в нативном необработанном виде. Антиген может также поглощаться В-клетками, которые могут процессировать его и представлять Т-клеткам.
Центроциты селектируются по их способности взаимодействовать с антигеном, удерживаемым дендритными клетками. Для центроцитов характерна высокая смертность путем апоптоза. Перекрестное связывание иммуноглобулиновых рецепторов со специфическим антигеном при одновременном проведении сигнала через рецептор CD40 отменяет апоптоз центроцитов. После получения сигналов через перечисленные рецепторы на центроцитах увеличивается экспрессия поверхностных иммуноглобулинов, и они приобретают характеристики В-клеток памяти.
Зародышевые центры сохраняются около трех недель после иммунизации, а после этого В-бласты памяти продолжают пролиферировать в фолликулах в течение месяцев, пока длится Т-зависимый антительный ответ. Эти клетки, вероятно, являются источником и плазматических клеток, и клеток памяти, необходимых для поддержания долгосрочной продукции антител и иммунологической памяти после первых трех недель Т-зависимого антительного ответа.
Описанные на нашем сайте реактивные лимфоидные клетки встречаются главным образом в лимфоидных тканях, их также можно увидеть в крови при инфекционном мононуклеозе и других вирусных инфекциях, а также у больных неходжкинскими лимфомами (в случае лейкемизации).
– Также рекомендуем “Гематология: Рециркуляция лимфоцитов”
Оглавление темы “Гематология”:
- Гематология: История изучения крови
- Концепция единства кровяной ткани. Кровь как мезенхимальная ткань
- Гематология: Эмбриология крови – кровяной ткани
- Гематология: Клетки и цитокины регулирующие развитие В-клеток крови
- Гематология: Развитие Т-лимфоцитов и его регуляция
- Гематология: Развитие лимфоцитов в лимфатических узлах и селезенке
- Гематология: Рециркуляция лимфоцитов
- Клиническое обследование пациента в гематологии: сбор анамнеза, объективный осмотр
- Морфологическое исследование клеток крови. Причины изменений
- Гематология: Лейкоцитарная формула в норме и при болезнях
Источник
Селезенка является одним из важнейших периферических органов иммунной системы. Неиммунная функция селезенки заключается в разрушении и утилизации погибающих или поврежденных форменных элементов крови, а также циркулирующих микроорганизмов. Селезенка — это мощный фильтр крови и барьер для чужеродных веществ на пути из магистрального сосуда (аорты) в систему воротной вены. Как периферический орган иммунной системы селезенка участвует в иммунном ответе на растворимые антигены, циркулирующие в крови, селекции, активации и развитии толерантности В-лимфоцитов.
Лимфатическая ткань селезенки имеет достаточно сложное анатомическое строение. Она представлена белой пульпой и лимфоидными элементами красной пульпы. Белая пульпа сосредоточена вдоль артериол селезенки и состоит из периартериолярных лимфоидных муфт, ассоциированных с ними лимфоидных фолликулов, эллипсоидных макрофагально-лимфо-идных муфт, располагающихся на концах ветвления артериальных сосудов, и венозных синусов, в которые впадают артериолы и истинные капилляры селезенки.
Схематично строение белой пульпы селезенки представлено на рисунке. В пределах периартериолярных лимфоидных муфт (ПЛМ) выделяют наружную и внутреннюю части. ПЛМ можно считать аналогом паракортикальной (тимусзависимой) зоны лимфатических узлов. Фолликулы представляют собой В-клеточные образования. Кроме того, в селезенке имеется особая популяция В-клеток, которые отграничивают белую пульпу (периартериолярные лимфоидные муфты, включающие первичные и вторичные фолликулы) от красной пульпы. Эта область получила название краевой, или маргинальной, зоны.
Т-клетки селезенки
В селезенке присутствуют только периферические (наивные и зрелые) Т-лимфоциты, прошедшие селекцию в тимусе. Под влиянием антигенного стимула эти клетки активируются подобно тому, как это происходит в лимфатических узлах.
В белой пульпе селезенки (в периартериолярных лимфоидных муфтах) CD4+ Т-клетки преобладают, CD8+ Т-клеток меньше, а в красной пульпе наблюдается обратное соотношение между этими популяциями.
Т-клетки, имеющие рецептор TCRyq, предпочтительно оседают в синусоидах селезенки, в то время как лимфоциты, несущие TCRab, заселяют главным образом периартериолярные лимфоидные муфты.
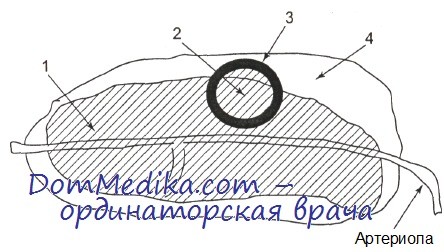
Строение белой пульпы селезенки; продольный срез вдоль артериолы. Показана периартериолярная лимфоидная муфта (1), включающая вторичный фолликул со светлым центром (2) и зоной мантии (3), а также маргинальную зону (4) по периферии лимфоидной ткани (схема)
В-клетки селезенки
В селезенке происходят процессы активации В-клеток в ходе первичных и вторичных иммунных ответов.
При формировании В-клеточного репертуара зрелые девственные В-лимфоциты поступают из костного мозга с током крови в Т-клеточные области периартериолярных лимфоидных муфт и далее в первичные фолликулы. В-клетки, специфичные в отношении аутологичных антигенов, не поступают в фолликулы, они задерживаются в наружной зоне ПЛМ и гибнут. Прошедшие фолликул В-клетки поступают в пул наивных В-лимфоцитов. Часть из них в дальнейшем активируется в селезенке, часть — в лимфатических узлах или других периферических лимфоидных органах.
Движение наивных В-клеток, поступающих из крови в селезенку, приостанавливается в наружной зоне ПЛМ. Это универсальное явление происходит после взаимодействия мембранных иммуноглобулинов В-лимфоцитов с антигеном. Биологический смысл процесса состоит в необходимости накопления активированных, пролиферирующих В-лимфоцитов в наружной зоне ПЛМ (маргинальная зона) в течение первых дней иммунного ответа на растворимые антигены белковой природы. Это позволяет многократно повысить вероятность встречи активированных наивных В-лимфоцитов с наивными Т-хелперными (CD4+) клетками той же антигенной специфичности, количество которых чрезвычайно мало.
Без Т-клеточной помощи, необходимой для реализации гуморального иммунного ответа на тимусзависимые антигены, активированные В-клетки погибают. При наличии Т-клеточной помощи наивные В-клетки поступают в фолликулы, где подвергаются соматическим гипермутациям и дифференцировке с изотипическим переключением в зародышевых центрах в ходе первичных иммунных ответов. При вторичных иммунных ответах В-клеток памяти на тимуезависимые антигены наблюдаются выраженная В-клеточная пролиферация и дифференцировка в плазматические клетки в пределах наружной зоны ПЛМ; фолликулярная В-клеточная пролиферация является несколько более слабой, чем при первичных ответах.
Антигенспецифические Т-клетки уничтожают в наружных зонах ПЛМ определенную часть В-лимфоцитов. Лиганд Fas-антигена, экспрессированный на этих Т-хелперах, включает программу гибели (апоптоз) в В-клетках с несвязанными антигенными рецепторами, а также в В-клетках, не имеющих нормального пути внутриклеточного проведения антигенного сигнала.
В тимуснезависимых иммунных ответах В-клетки способны дифференцироваться в плазмоциты без Т-клеточной помощи. При ответе на TI-антигены (ЛПС) происходят выраженная антигенспецифическая В-клеточная пролиферация и плазмоклеточная дифференцировка в наружной зоне ПЛМ и в красной пульпе; фолликулярная В-клеточная пролиферация умеренная. Считают, что именно поликлональные активаторы типа ТЫ, а также аутологичные антигены ведут к индукции CD5 на В-лимфоцитах.
CD5+ В-клетки обычно не проходят через светлый центр и не подвергаются изотипическому переключению. Интересно существование некоторого сходства CD5-лимфоцитов с толерантными аутореактивными В-клетками, которые в наружных зонах ПЛМ подвергаются абортивной активации, препятствующей их дальнейшей миграции в фолликул.
В TI-2-ответах (фиколл) большинство пролиферирующих В-клеток в наружной зоне ПЛМ дифференцируется в плазматические клетки. В селезенке существует особая область, окружающая периартериолярные лимфоидные муфты, так называемая краевая (маргинальная) зона. Клетки этой зоны являются нормальным эквивалентом клеток, из которых возникают лимфомы маргинальной зоны, или лимфоцитомы селезенки. В маргинальной зоне селезенки реализуются В-клеточные иммунные ответы на тимуснезависимые антигены, циркулирующие в периферической крови. В-клетки маргинальной зоны имеют специфические морфологические и иммунологические черты. На мембране В-лимфоцитов маргинальной зоны селезенки экспрессированы IgM, но отсутствуют IgD. Эти клетки не являются рециркулирующими, специализированы к иммунному ответу на тимуснезависимые углеводные антигены. Вместе с тем эти лимфоциты происходят из пула рециркулирующих В-клеток, возвратившихся в маргинальную зону селезенки (хоминг).
Клетки маргинальной зоны расположены в сети первичных кровяных синусоидов селезенки, что позволяет им взаимодействовать с антигенами, переносимыми кровью. Популяция клеток маргинальной зоны не столь гомогенна, как принято считать. Сюда входят В-клетки памяти, генерированные как в ходе Т-зависимых, так и Т-независимых (I тип) антительных ответов, а также девственные В-клетки, которые еще не подверглись пролиферации, зависящей от присутствия антигена. Клетки маргинальной зоны способны in vivo отвечать на все типы антигенов: Т-зависимые, Т-независимые I и II типов. Характеристики этих клеток указывают на то, что они покоятся в фазе G1, а не G0.
Ответ in vivo на TI-2-антигены (но не на Til-антигены) сильно повреждается при спленэктомии, что подтверждает отсутствие способности к рециркуляции клеток маргинальной зоны селезенки.
Их иммунофенотип сходен с иммунофенотипом большинства рециркулирующих В-клеток, но есть и некоторые отличия. Обобщенно иммунофенотипические черты В-клеток маргинальной зоны селезенки можно представить следующим образом: sIgM+ slgD- CD21+ CD35+ CD23- CD76+/- CD75-/+ CD78-2C9-/+ CD22+ CD37+ CD24- CD19+/- CD20+/- CD44+/-CD10.
В последние годы получены новые данные относительно роли селезенки в формировании репертуара длительно живущих наивных В-лимфоцитов. Из числа ежедневно продуцируемых костным мозгом ~ 1010 девственных В-лимфоцитов примерно 10% переходят в длительно живущий пул. Девственные В-лим-фоциты IgM+ IgD+ CD23- CD5- поступают первоначально в селезенку. Через концевые ветви центральных артериол и синусы маргинальной зоны они попадают в наружную зону ПЛМ. Далее небольшая часть В-клеток поступает в фолликулы. Именно эти клетки переходят в долгоживущий рециркулирующий пул. Положительная селекция В-лимфоцитов осуществляется на основании экспрессии IgVH-генов в ответ на низкие дозы экзогенных антигенов или на элементы идиотипической сети (например, сывороточные иммуноглобулины). В процессе перехода девственных клеток в наивные длительно живущие рециркулирующие лимфоциты не происходит соматических гипермутаций и переключения классов иммуноглобулинов. Пока не совсем понятно, в чем состоят иммунофенотипические различия между девственными и наивными В-лимфоцитами. Девственные В-клетки, не подвергнувшиеся положительной селекции и не поступившие в фолликулы, погибают в наружной зоне ПЛМ.
– Читать “Костный мозг как лимфоидный орган. Лимфопоэз в костном мозге”
Оглавление темы “Клетки крови”:
- Онтогенез Т-лимфоцитов – образование
- Созревание (онтогенез) NK-клеток
- Созревание (онтогенез) лимфоидных дендритных клеток Лангерганса
- Рециркуляция лимфоцитов в организме
- Лимфатические узлы – формирование Т-лимфоцитов
- Функции Т-лимфоцитов лимфатических узлов
- В-лимфоциты лимфатических узлов – созревание, функции
- Внутрифолликулярная дифференцировка В-лимфоцитов
- Селезенка – строение, формирование Т- и В-лимфоцитов
- Костный мозг как лимфоидный орган. Лимфопоэз в костном мозге
Источник
Оглавление темы “Селезенка. Лимфатические узлы. Стадии иммунного ответа. Формы иммунного ответа. Воспаление. Механизмы, контролирующие иммунную систему.”: Селезенка. Функции селезенки. Лимфатические узлы. Функции лимфатических узлов.Селезенка выполняет функции фильтра крови, удаляющего из крови попадающие туда чужеродные частицы и молекулы, а также состарившиеся эритроциты.
В селезенке имеются Т- и В-зоны, заселенные преимущественно Т- или В-лимфоцитами. В качестве одного из периферических органов иммунной системы селезенка является местом: Селезенка является местом специфического иммунного ответа на антигены, циркулирующие в крови, а в лимфатических узлах разыгрываются процессы специфического иммунного ответе на антигены, попадающие через лимфу (рис. 8.5). Лимфатические узлыЛимфатические узлы функционируют в качестве своеобразных фильтров лимфы, задерживая микроорганизмы, попавшие в лимфу. В качестве периферических органов иммунной системы лимфатические узлы являются местом:
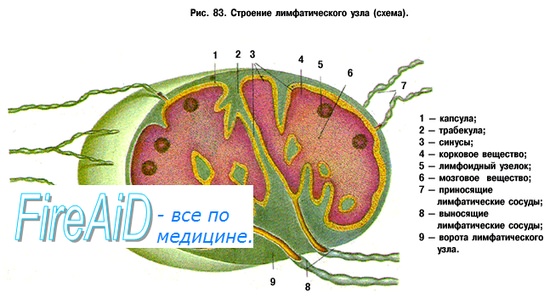
Один лимфатический узел имеет массу около 1 г. Каждый час из лимфоузла выходит в лимфу количество лимфоцитов, эквивалентное его утроенной массе. Большая часть (90 %) клеток в этой эфферентной лимфе представляют собой лимфоциты, покинувшие кровяное русло на территории этого лимфатического узла. Среди клеток лимфатического узла около 10 % составляют макрофаги и около 1 % — дендритные клетки. В кортикальном (наружном) слое лимфатического узла преобладают скопления лимфоцитов. В мозговом (внутреннем) слое лимфатического узла лимфоциты содержатся в сочетании с макрофагами (около 10 %) и дендритными клетками (около 1 %), которые выполняют функцию представления антигенов Т-лимфоцитам и продуцируют цитокины. В пределах лимфатического узла происходит свободная циркуляции лимфоцитов между лимфой, кровью и лимфоидной тканью. При развитии специфического иммунного ответа в мозговом слое лимфатических узлов накапливаются образовавшиеся из В-лимфоцитов плазматические клетки, продуцирующие и секретирующие соответствующие антигену по специфичности антитела —иммуноглобулины. Строго определенное взаимное расположение Т- и В-лимфоцитов, макрофагов, дендритных клеток в лимфатических узлах обеспечивает последовательное развитие стадий иммунного ответа: представления антигена, его распознавания Т-клеточными рецепторами, активации процессов пролиферации и дифференцировки (созревания) лимфоцитов. В результате в лимфатических узлах формируются зрелые Т-лимфоциты, которые принимают участие в защитных реакциях. Там же формируются клоны долгоживущих Т- и В-лимфоцитов, обеспечивающих поддержание иммунологической памяти о встрече с конкретным антигеном. – Также рекомендуем “Мукозно-ассоциированная лимфоидная ткань. Лимфоидная ткань слизистых оболочек.” |

Источник