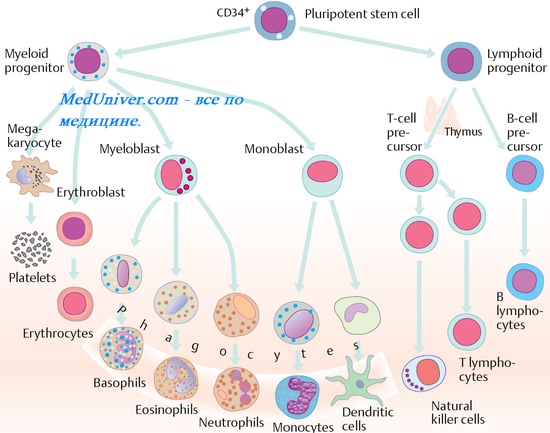
Этапы созревания в лимфоцитов

Оглавление темы “СD8 лимфоциты. Антиген ( Аг ) представляющие клетки. Классификация антигенов ( Аг ).”:
1. Рецептор Т-клеток. Активация Т-лимфоцитов. Субпопуляции Т-лимфоцитов. СD4+ лимфоциты.
2. СD8 лимфоциты. Т супрессоры. Цитотоксические Т-лимфоциты ( ЦТЛ ). Т-киллеры. Цитотоксический эффект Т-киллеров. Т-клетки памяти.
3. В-лимфоциты. В-клетки. Созревание В-лимфоцитов. Продолжительность жизни B-лимфоцитов. Маркеры В-клеток ( В-лимфоцитов ).
4. Естественные киллеры ( NK-клетки ). Перфорины. Функция естественных киллеров. Задачи NK-клеток.
5. Антиген ( Аг ) представляющие клетки. Иммунные реакции. Классификация имунных реакций. Антигены. Свойства антигенов. Структура антигенов ( Аг ). Валентность Аг.
6. Классификация антигенов ( Аг ). Иммуногены. Гаптены. Непреципитирующие гаптены. Преципитирующие гаптены. Полугаптены. Проантигены.
7. Адъюванты. Адъювант Фройнда. Толерогены. Природа антигенов (Аг). Происхождение антигенов. Молекулярная масса антигенов (Аг).
8. Специфичность антигенов (Аг). Видовые антигены. Групповые антигены (Аг). Гетерогенные антигены (Аг). Антиген Форссмана. Аллоантигены ( изоантигены ).
9. Патологические антигены. Аутоантигены. Врождённые аутоантигены. Приобретённые аутоантигены. Суперантигены.
10. Антигены ( Аг ) микроорганизмов. Антигены бактерий. Капсульные антигены ( К-антигены ( Аг )). Соматические антигены ( O-антигены ( Аг )). Жгутиковые антигены ( H-антигены ( Аг )). Vi-Ar ( Антиген вирулентности ).
В-лимфоциты. В-клетки. Созревание В-лимфоцитов. Продолжительность жизни B-лимфоцитов. Маркеры В-клеток ( В-лимфоцитов ).
Известны субпопуляции В-клеток: предшественники антителообразующих (плазматических) клеток и В-клетки памяти (эффекторы вторичных иммунных реакций). Доминирующую субпопуляцию составляют предшественники антителообразующих клеток, дифференцирующиеся после антигенной стимуляции в плазматические клетки (плазмоциты), синтезирующие Ig.
Созревание В-лимфоцитов
Из костного мозга пре-В-клетки мигрируют в тимуснезависимые зоны лимфоидных органов. Так, в физиологических условиях в селезёнке В-лимфоциты располагаются в краевой зоне белой пульпы, в лимфатических узлах — в наружной зоне кортикального слоя, где они формируют зародышевые центры фолликулов. Сигналы, определяющие судьбу и дифференцировку этих иммунокомпетентных клеток, поступают из красного костного мозга, стромальных клеток и других клеток иммунной системы.
На периферии (вне костного мозга) В-лимфоциты приобретают характерные для них поверхностно-клеточные маркёры. Продолжительность жизни В-лимфоцитов различна — от многих лет (В-клетки памяти) до нескольких недель (клоны плазматических клеток).
После антигенной стимуляции В-лимфоциты дифференцируются в плазматические клетки (интенсивно синтезирующие и секретирующие AT) и В-клетки памяти. Плазматические клетки синтезируют Ig того же класса, что и мембранный Ig В-лимфоцита-предшественника.
Маркеры В-клеток ( В-лимфоцитов )
Основные маркёры В-лимфоцитов — мембранные Ig при этом клетки одного клона (быстро формирующегося в результате серии последовательных делений потом-ства одной В-клетки ) экспрессируют молекулы Ig, специфически связывающие только один эпитоп Аг. Такие клетки синтезируют моноклональные AT, способные распознавать и связывать только один Аг. Аг-связывающий участок мембранного Ig В-лимфоцита играет роль клеточного Аг-распознающего рецептора.
Помимо мембранных Ig, В-лимфоцит несёт другие маркёры; рецепторы Fc-фрагмента Ig, CD10 {на незрелых В-клетках), CD19, CD20, CD21, CD22, CD23 (вероятно, участвуют в клеточной активации), рецепторы к С3b и C3d, молекулы МНС классов I и И.
– Также рекомендуем “Естественные киллеры ( NK-клетки ). Перфорины. Функция естественных киллеров. Задачи NK-клеток.”
Источник
– Про-В-клетка экспрессирует ферменты, необходимые для реаранжировки генов и соединительного разнообразия, однако ни тяжелые, ни легкие цепи при этом не экспрессируются.
– Пре-В-клетка экспрессирует тяжелые μ-цепи в цитоплазме. На клеточной поверхности тяжелые цепи соединяются с суррогатными легкими цепями для образования рецепторов пре-В-клетки.
– Незрелая В-клетка синтезирует легкие цепи и, следовательно, может экспрессировать молекулы антител на клеточной поверхности. При взаимодействии с антигеном на этом этапе может произойти отрицательный отбор.
– На стадии переходных (транзиторных) В-клетки постепенно становятся нечувствительны к негативному отбору и приобретают статус иммунокомпетентных
– Зрелая В-клетка экспрессирует как иммуноглобулин М, так и иммуноглобулин D и способна компетентно реагировать на антиген.
Перед превращением в зрелые В-лимфоциты клетки проходят ряд этапов развития, что приводит к последовательному формированию тяжелых цепей, легких цепей и целых молекул антител. В определенные моменты этого процесса происходит их отбор с удалением клеток с бесцельной перестройкой генов или формированием аутореактивных антител, а также с подачей сигналов выживаемости для клеток, которые могут производить потенциально применимые антитела.
Процесс развития В-лимфоцитов делится на несколько этапов, каждый из которых характеризуется определенными событиями, при этом клетки на разных этапах отличаются поверхностными клеточными маркерами и экспрессией генов иммуноглобулинов.
Гемопоэтические стволовые клетки костного мозга и эмбриональной печени, из которых происходят В-клетки, изначально являются плюрипотентными. Стволовые клетки, которые исходно развивались по лимфоцитарному пути, могут стать общими предшественниками лимфоцитов, которые в свою очередь превращаются в В-, Т- или NK-клетки. В-клетки, происходящие из печени плода, главным образом представлены В1-клетками, в то время как В-клетки костномозгового происхождения исходно представляют собой фолликулярные В-лимфоциты.
Клетки и молекулы, расположенные внеклеточно, в стромальном микроокружении вырабатывают сигналы, приводящие к дифференцировке лимфоцитов. Индукция регуляторов транскрипции EBF, Е2А и Рах-5 приводит к экспрессии белков, критически значимых для развития В-лимфоцитов. Дополнительный контроль дифференцировки В-клеток обеспечивается посттранскрипционной регуляцией мРНК РНК-связывающими белками и микроРНК.
Самый ранний предшественник В-лимфоцитов называется пре-В клеткой. На этом этапе клетка вырабатывает ген, активирующий рекомбинацию (ГАР) и терминальные ДНК-трансферазы (ТдТ), которые в последующем потребуются для соматической рекомбинации и переноса нуклеозидов, обеспечивающих соединительное разнообразие, соответственно. На этапе пре-В-клетки происходит ограниченная соматическая рекомбинация, а иммуноглобулины еще не экспрессируются.
Следующим этапом созревания В-клетки является пре-В-клетка. На этой стадии происходит синтез тяжелой цитоплазматической μ-цепи. Поскольку легкие цепи в этот момент еще не экспрессируются, поверхностный иммуноглобулин на данном этапе не присутствует. Некоторые из p-тяжелых цепей соединяются с постоянными молекулами, которые называются суррогатами легких цепей, а также с белками иммуноглобулинов α и β, обеспечивающими передачу сигнала с образованием комплексов, которые носят название пре-В-рецепторы.
На данном этапе селектируются клетки, которые синтезировали тяжелые цепи, способные к формированию части пре-В-клеточного рецептора, поскольку последний вырабатывает сигналы, необходимые для выживания, размножения и созревания клеток.
Образование легких цепей происходит на следующей стадии созревания В-клеток, когда появляется незрелая В-клетка. В результате соединения легких цепей с тяжелыми p-цепями возникает молекула IgM, которая может экспрессироваться на клеточной поверхности. Несмотря на то что присутствие В-клеточного рецепторного комплекса мотивирует способность клетки распознавать специфические антигены, на данной стадии подобное распознавание не приводит к пролиферации или дифференцировке. Напротив, клетки могут подвергнуться отрицательному отбору при встрече с антигеном.
Незрелые В-клетки, распознающие аутоантиген, могут ликвидироваться путем апоптоза или же пройти через процесс рецепторного редактирования, в ходе которого приобретается новая специфичность, не являющаяся аутоспецифичностью.
Выход незрелых В-клеток из костного мозга в селезенку знаменует начало новой стадии появление переходных В-лимфоцитов. Переходные клетки постепенно начинают экспрессировать на своей поверхности IgD, CD21 и CD23 и становятся более иммунокомпетентными. Альтернативный сплайсинг РНК обеспечивает синхронную экспрессию IgM и IgD. В начале данного этапа перекрестная активация В-клеточного рецептора ведет к негативному отбору. По мере дальнейшего созревания переходные клетки становятся чувствительными к действию Т-хелперов и, в то же время теряют чувствительность к негативному отбору.
Зрелая В-клетка вырабатывает IgM и IgD и может полноценно отвечать на действие антигена. Такая клетка считается наивной, потому что она еще не активировалась антигеном. Большинство зрелых В-лимфоцитов циркулируют через периферическую лимфатическую систему (селезенку, лимфатические узлы, лимфоидную ткань слизистых оболочек). Они носят название фолликулярных В-клеток или рециркулирующих В-клеток. В-лимфоциты привлекаются в фолликулы хемокином СХСL13, секретируемым фолликулярными дендритными клетками. В-лимфоциты выживают в фолликулах благодаря цитокину BAFF (В-cell activating factor—фактор, активирующий В-клетки), также известному как BLyS (В lymphocyte stimulator—стимулятор В-лимфоцитов). Небольшая часть зрелых В-клеток возвращается в краевую зону селезенки и остается там.
Взаимодействие зрелых наивных В-клеток с антигеном приводит к активации В-клеток, их пролиферации и дифференцировке. При этом отдельная группа В-клеток трансформируется в В-клетки памяти, которые активируются позже в случае последующей встречи с антигеном. Часть других В-клеток дифференцируется в клетки, постепенно уменьшающие количество производимых ими мембраносвязанных Ig, увеличивая при этом выработку секретированных форм Ig.
Терминально дифференцированные В-клетки, предназначенные для выработки секретированных Ig, представляют собой плазматические клетки, имеющие обширную шероховатую эндоплазматическую сеть, что также соответствует функции таких клеток служить фабриками антител.
– Вернуться в оглавление раздела “дерматология.”
Оглавление темы “Гуморальный иммунитет и антитела при болезнях кожи.”:
- Гуморальный иммунитет и структура антител
- Виды иммуноглобулинов и их функции
- Механизмы обеспечивающие разнообразие антител (иммуноглобулинов)
- Механизмы и этапы созревания В-лимфоцитов
Источник
Часть лимфоидных стволовых клеток мигрирует в вилочковую
железу (тимус) и созревает там до Т-лимфоцитов; другая их часть остается в
костном мозге и созревает до В-лимфоцитов. На пути созревания (дифференцировки)
в центральных органах иммунитета стволовая клетка проходит несколько этапов.
Эти этапы созревания Т- и В-лимфоцитов происходят без участия антигена и
поэтому получили название антиген независимой дифференцировки.
Первый этап – это появление на стволовой клетке структур, указывающих,
по какому пути дифференцировки (Т- или В-лимфоцитов) пойдет ее развитие; он
происходит еще в костном мозге.
Ранний предшественник В-лимфоцитов имеет на своей мембране так
называемую суррогатную L-цепь молекулы иммуноглобулина, а ранний предшественник
Т-лимфоцитов – гликопротеин с молекулярной массой 3,3*104 D (ГП-33), который
впоследствии ассоциируется с бета-цепью Т-клеточного антигенораспознающего
рецептора.
Указанные структуры, определяющие дальнейшую судьбу
гемопоэтической стволовой клетки, играют важную роль: 1) задают путь
дифференцировки Т- и В-лимфоцитов; 2) передают сигнал ранним предшественникам
Т- и В-лимфоцитов, по которому начинается их пролиферация.
Например, на этапе появления суррогатной L-цепи и ГП-33
таких клеток в костном мозге и тимусе у мышей насчитывается по 3-5*106. Однако
уже через 4 недели их количество увеличивается в 10-100 раз и составляет 5*107
– 108 клеток. Таким образом, наличие суррогатной L-цепи и ГП-33 является тем
инструментом, с помощью которого количество пре-В- и пре-Т-клеток достигает
необходимого уровня.
Второй этап дифференцировки лимфоцитов- это появление
незрелых предшественников В- и Т-лимфоцитов.
Он определяется появлением на мембране В- и Т-лимфоцитов
антигенраспознающих рецепторов, с помощью которых В- и Т-лимфоциты после этого
этапа дифференцировки приобретают способность распознавать антигены.
Для В-лимфоцита антигенраспознающим рецептором (ВАГРР)
является мембранный IgM, а для Т-лимфоцита (ТАГРР) – специальная димерная молекула,
имеющая альфа- и-бетацепи, и относящаяся к супер семейству иммуноглобулинов
(подробно об антигенраспознающих рецеп-торах будет сказано в разделах о Т- и
В-лимфоцитах).
Между I и II этапом есть еще несколько стадий дифференцировки,
о которых здесь не упоминается.Таким образом, появление на поверхности ранних
предшественников лимфоидных клеток определенных структур (рецепторов) служит
тем сигналом, который позволяет клеткам дифференцироваться в специфическую
линию лимфоцитов. Имеющие такие рецепторы клетки-предшественники мигрируют в
определенную область центральных иммунных органов, а затем взаимодействуют со
специфическим микроокружением, необходимым для того, чтобы в этой области
произошла дифференцировка данной клетки. В ответ на контакт с
клеткой-предшественником, в стромальных клетках локального микроокружения в
свою очередь развиваются молекулярные процессы, которые направлены на
“обучение” (коммитирование) клеток-предшественников для дальнейшей их
дифференцировки в отдельно взятую линию.
Показано, что клетки-предшественники в тесном контакте с их
стромальным клеточным ми-кроокружением сохраняют очередную стадию
дифференцировки до тех пор, пока имеется достаточно пространства в соответствующей зоне (компартменте)
центрального органа иммунитета (тимуса или костного мозга), и пока
соответствующий цитокин, ответственный за пролифера-цию и дифференцировку
продуцируется в данной зоне.
Таким образом, отсутствие контакта со стромальными клетками
и потеря сигнала через ци-токиновый рецептор являются теми условиями, при
которых клетка-предшественник переходит к дальнейшей стадии дифференцировки.
Это позволяет ей осуществить новый контакт в следующем, новом месте
микроокружения.
Источник
Часть лимфоидных стволовых клеток мигрирует в вилочковую железу (тимус) и созревает там до Т-лимфоцитов; другая их часть остается в костном мозге и созревает до В-лимфоцитов. На пути созревания (дифференцировки) в центральных органах иммунитета стволовая клетка проходит несколько этапов. Эти этапы созревания Т- и В-лим- фоцитов происходят без участия антигена и поэтому получили название антигеннезависимойдифференцировки.
Первый этап —это появление на стволовой клетке структур, указывающих, по какому пути дифференцировки (Т- или В-лимфоцитов) пойдет ее развитие; он происходит еще в костном мозге.
Ранний предшественник В-лимфоцитов имеет на своей мембране так называемую суррогатную L-цепь молекулы иммуноглобулина, а ранний предшественник Т-лимфоцитов — гликопротеин с молекулярной массой 3,3 • 104 D (ГП-33), который впоследствии ассоциируется с бета-цепью Т-клеточного антигенраспознающего рецептора.
Указанные структуры, определяющие дальнейшую судьбу гемопоэтической стволовой клетки, играют важную роль: 1) задают путь дифференцировки Т- и В-лимфоцитов; 2) передают сигнал ранним предшественникам Т- и В-лимфоцитов, по которому начинается их пролиферация.
Например, на этапе появления суррогатной L-цепи и ГП-33 таких клеток в костном мозге и тимусе у мышей насчитывается по 3—5 • 106. Однако уже через 4 недели их количество увеличивается в 10—100 раз и составляет 5 НО7— 108 клеток. Таким образом, наличие суррогатной L-цепи и ГП-33 является тем инструментом, с помощью которого количество пре-В- и пре-Т-клеток достигает необходимого уровня.
Второй этап дифференцировки лимфоцитов —это появление незрелых предшественников В- и Т-лимфоцитов.
Он определяется появлением на мембране В- и Т-лимфоцитов ан- тигенраспознающих рецепторов, с помощью которых В- и Т-лимфо- циты после этого этапа дифференцировки приобретают способность распознавать антигены.
Для В-лимфоцита антигенраспознающим рецептором (ВАГРР) является мембранный IgM, а для Т-лимфоцита (ТАГРР) — специальная димерная молекула, имеющая альфа- и бета-цепи, и относящаяся к суперсемейству иммуноглобулинов (подробно об антигенраспознающих рецепторах будет сказано в разделах о Т- и В-лимфоцитах).
Между I и II этапом есть еще несколько стадий дифференцировки, о которых здесь не упоминается. ¦
Таким образом, появление на поверхности ранних предшественников лимфоидных клеток определенных структур (рецепторов) служит тем сигналом, который позволяет клеткам дифференцироваться в . специфическую линию лимфоцитов. Имеющие такие рецепторы клетки- предшественники мигрируют в определенную область центральных иммунных органов, а затем взаимодействуют со специфическим микроокружением, необходимым для того, чтобы в этой области произошла дифференцировка данной клетки. В ответ на контакт с клеткой- предшественником, в стромальных клетках локального микроокружения в свою очередь развиваются молекулярные процессы, которые направлены на “обучение” (коммитирование) клеток-предшественни- ков для дальнейшей их дифференцировки в отдельно взятую линию.
Показано, что клетки-предшественники в тесном контакте с их стромальным клеточным микроокружением сохраняют очередную стадию дифференцировки до тех пор, пока имеется достаточно пространства в соответствующей зоне (компартменте) центрального органа иммунитета (тимуса или костного мозга), и пока соответствующий цитокин, ответственный за пролиферацию и дифференцировку продуцируется в данной зоне.
. Таким образом, отсутствие контакта со стромальными клетками и потеря сигнала через цитокиновый рецептор являются теми условиями, при которых клетка-предшественник переходит к дальнейшей стадии дифференцировки. Это позволяет, ей осуществить новый контакт в следующем, новом месте микроокружения.
Источник