Хоминг рецепторы в рециркуляции лимфоцитов

Способность клеток находить «свое место» в организме называют хомингом. Существует две группы механизмов, задействованных в данном процессе, — контактные механизмы, обеспечиваемые молекулами адгезии, находящимися на поверхности лимфоцитов, и хемотаксис, определяющий направленность движения клеток, зависящую от наличия на их поверхности хемокиновых рецепторов.
Попав в сосудистую сеть лимфоидного органа, лимфоциты реагируют на хемокины, вырабатываемые клетками эндотелия в специализированном для миграции лимфоцитов отделе сосудов — посткапиллярных венулах. Только в посткапиллярных венулах лимфоидных (но не других) органов есть высокий эндотелий, образованный спонтанно активированными клетками (аналогичный высокий эндотелий может быть индуцирован в других органах только под влиянием провоспалительных факторов). Клетки этого эндотелия секретируют в-хемокин ССL19, распознаваемый рецептором CCR7. Этот рецептор экспрессируют на поверхности наивные лимфоциты как Т-, так и В-класса. Таким образом, указанные эндотелиальные клетки способны привлекать как Т-, так и В-лимфоциты. Далее происходит трансмиграция лимфоцитов по механизму, аналогичному миграции лейкоцитов в очаг воспаления (отличия касаются конкретных молекул адгезии, определяющих направление миграции — рис. 3.74). Ключевую роль при этом играет начальный момент миграции, обусловленный взаимодействием мембранного L-cелектина (СD62L) лимфоцитов с адрессинами (GlyCAM, CD34, PSGL-1) эндотелиальных клеток. Именно наличие CD62L на клеточной мембране определяет способность наивных лимфоцитов мигрировать в лимфатические узлы и пейеровы бляшки, в связи с чем CD62L называют рецептором хоминга. Это слабое взаимодействие обусловливает феномен качения (перекатывания лимфоцита вдоль сосудистой стенки). Вслед за этим устанавливается более прочное взаимодействие между в2-интегрином лимфоцитов LFA-1 и его рецептором на эндотелиальной клетке — ICAM-1. Взаимодействие усиливается благодаря «активации»в2-интегрина (т.е. изменения конформации молекулы, в результате которого повышается его сродство к рецептору. Это обусловливает прочное прилипание лимфоцита к сосудистой стенке.
Затем под влиянием хемотаксических сигналов, поступающих из ткани лимфоидного органа, лимфоцит начинает мигрировать из сосуда между эндотелиальными клетками.
Вышедшие из сосуда клетки попадают в примыкающие к посткапиллярным венулам тимусзависимые зоны, стромальные и дендритные клетки которых секретируют хемокины CCL19 (ELC) и ССL21 (SLC), распознаваемые рецептором CCR7 на лимфоцитах. До этого момента Ти В-клетки перемещаются в одном и том же направлении. После того как лимфоцит попадает во внутреннюю среду органа, их пути расходятся. Т-клетки остаются в тимусзависимой зоне (в паракортексе лимфатических узлов, параартериальной муфте белой пульпы селезенки и Т-зонах пейеровых бляшек). В-клетки реагируют на хемокин CXCR13 (BLC), к которому они экспрессирут рецептор СХСR5. Этот хемокин секретируют стромальные клетки фолликулов, что и определяет направление миграции В-лимфоцитов. В-лимфоциты, не несущие СХСR5, не поступают в фолликулы и локализуются в прилежащей к фолликулу зоне. Механизмы, определяющие дальнейшее перемещение лимфоцитов из органа в эфферентную лимфу, пока не установлены.
В процессе рециркуляции отсутствует «привязка» хоминга к конкретному лимфатическому узлу или пейеровой бляшке. Так, лимфоцит, покинувший шейный лимфоузел, может затем проникнуть, например, в паховый лимфатический узел или в пейерову бляшку. Принципиально лишь то, что клетки не выходят из системы вторичных лимфоидных органов. Степень специфичности хоминга минимальна в селезенке, так как лимфоциты проникают в белую пульпу при излиянии крови в краевой зоне. Только последующее перемещение в Ти В-зоны осуществляется по описанному выше механизму.
Все сказанное выше относится к наивным лимфоцитам. После активации лимфоцитов антигеном экспрессия молекул адгезии и хемокиновых рецепторов на их поверхности существенно изменяется. Это определяет иные пути миграции клеток после контакта с антигеном. Активированные лимфоциты и клетки памяти очень слабо экспрессируют L-селектин, что обусловливает низкую вероятность их попадания в лимфатические узлы. В то же время они приобретают способность мигрировать в очаги воспаления, расположенные в нелимфоидных органах, а также в барьерные ткани .Пути миграции активированных лимфоцитов и клеток ткани памяти будут специально рассмотрены далее .Благодаря различной экспрессии рецепторов хоминга в разных тканях формируется несколько относительно автономных путей рециркуляции. Пути рециркуляции наивных лимфоцитов и клеток памяти могут пересекаться. Так, и наивные клетки, и клетки памяти, активированные в кишечнике, попадают в брыжеечные лимфатические узлы. Интенсивность рециркуляции Т-лимфоцтов существенно выше, чем В-лимфоцитов. У человека через каждый лимфатический узел за сутки проходит 0,3Ч1011, через селезенку — 2,5Ч1011, а через кровь — 5Ч1011лимфоцитов.
Длительность пребывания Т-клеток в кровотоке за 1 цикл рециркуляции — около 30 мин. Благодаря рециркуляции происходит постоянное «перемешивание» лимфоидных клеток с сохранением описанного выше порядка: наивные клетки, как правило, не покидают лимфоидной ткани, тогда как клетки памяти, наоборот, часто выходят за ее пределы, мигрируя, напри- мер, в эпителиальные органы и ткани, а также в очаги воспаления. Внутри лимфоидных органов клетки также занимают соответствующие места. Упорядоченность локализации лимфоцитов различных типов очень важна для развития иммунного ответа
Источник
Лимфоидные органы — динамичные образования: лимфоциты непрерывно перемещаются в стромальном каркасе, покидают лимфоидные органы и вновь возвращаются в них. Этот круговорот лимфоцитов обозначают термином «рециркуляция». В процессе рециркуляции лимфоциты покидают лимфоидные органы с эфферентной лимфой (из селезенки они выходят с кровью). Через грудной лимфатический проток лимфоциты проникают в кровяное русло. Затем через посткапиллярные венулы клетки вновь проникают в лимфоидный орган (тот же или другой), внутри которого перемещаются в специфические места их локализации. Затем клетки вновь покидают орган и выходят в циркуляторное русло и т.д. В нелимфоидных органах лимфоциты практически не циркулируют.
Из сказанного следует, что выявляемые в крови лимфоциты находятся в ней недолго (примерно 30 мин), но в течение суток многократно (4—5 раз) покидают ее и вновь возвращаются. Таким образом, несмотря на то, что в крови лимфоциты составляют всего около 0,5% от общего числа клеток, их состояние достаточно точно отражает состояние популяции лимфоцитов в целом организме. Это очень существенно, поскольку именно лимфоциты крови человека служат наиболее обычным и доступным объектом изучения как в экспериментальных, так и в клинико-иммунологических исследованиях. Субпопуляционный состав лимфоцитов крови человека представлен в табл. 3.23.
Таблица 3.23. Нормальные показатели содержания в крови взрослых людей основных популяций и субпопуляций лимфоцитов (по С.В. Хайдукову, 2008)
| Клетки | Относительное содержание*, % | Число клеток в 1 л (М+m) |
| Лимфоциты (CD45+)** | 32+4 | 1,363—2,808х109 |
| В-клетки ^D3-CD19+) | 12+5 | 0,111—0,376х109 |
| Т-клетки (CD3+CD19-) | 73+12 | 0,946-2,079х109 |
| арТ-клетки (CD3+apTCR+) | 70,5+9,7 | 0,022—0,115х109 |
| yST-клетки (CD3+ySTCR+) | 4,6+2,8 | 0,924—1,964х109 |
| Т-хелперы (CD3+CD4+) | 45+10 | 0,576—1,336х109 |
| Т-киллеры (CD3+CD8+) | 27+8 | 0,372—0,974х109 |
| Регулятоные T-клетки (CD3+CD4+CD25hi) | 3,7+2,05 | 0,009—0,078х 109 |
| NKT-клетки (CD3+CD56+CD16+) | 3+3 | 0,007—0,165х109 |
| Т-хелперы наивные (CD3+CD4+CD45RA+) | 30+10 | 0,272—1,123х109 |
| Т-хелперы активированные/памяти (CD3+CD4+CD45R0+CD29+) | 15+10 | 0,068—0,702х109 |
| NK-клетки (CD45+CD56+СD16+) | 13+5 | 0,123—0,369х109 |
* Указано среднее арифметическое со стандартной ошибкой.
** Относительные и абсолютные количества лимфоцитов определяли от общего количества лейкоцитов.
Относительные и абсолютные количества субпопуляций Т-, В- и NK-клеток определяли от общего количества лимфоцитов.
Способность клеток находить «свое место» в организме называют хомингом. Существует две группы механизмов, задействованных в данном процессе, — контактные механизмы, обеспечиваемые молекулами адгезии, находящимися на поверхности лимфоцитов, и хемотаксис, определяющий направленность движения клеток, зависящую от наличия на их поверхности хемокиновых рецепторов.
Попав в сосудистую сеть лимфоидного органа, лимфоциты реагируют на хемокины, вырабатываемые клетками эндотелия в специализированном для миграции лимфоцитов отделе сосудов — посткапиллярных венулах.
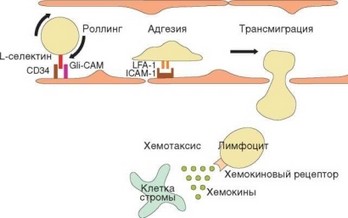
Рис. 3.74. Схема транссосудистой миграции лимфоцитов. Отражены основные стадии эмиграции лимфоцитов через высокий эндотелий посткапиллярных венул лимфоидных органов. Указаны молекулы адгезии, важные для осуществления этого процесса; отражена роль хемокинов в привлечении клеток
Только в посткапиллярных венулах лимфоидных (но не других) органов есть высокий эндотелий, образованный спонтанно активированными клетками (аналогичный высокий эндотелий может быть индуцирован в других органах только под влиянием провоспалительных факторов). Клетки этого эндотелия секретируют р-хемокин ССЫ9, распознаваемый рецептором CCR7. Этот рецептор экспрессируют на поверхности наивные лимфоциты как Т-, так и В-класса. Таким образом, указанные эндотелиальные клетки способны привлекать как Т-, так и В-лимфоциты.
Далее происходит трансмиграция лимфоцитов по механизму, аналогичному миграции лейкоцитов в очаг воспаления (отличия касаются конкретных молекул адгезии, определяющих направление миграции — рис. 3.74). Ключевую роль при этом играет начальный момент миграции, обусловленный взаимодействием мембранного L-cелектина (СD62L) лимфоцитов с адрессинами (GlyCAM, CD34, PSGL-1) эндотелиальных клеток. Именно наличие CD62L на клеточной мембране определяет способность наивных лимфоцитов мигрировать в лимфатические узлы и пейеровы бляшки, в связи с чем CD62L называют рецептором хоминга. Это слабое взаимодействие обусловливает феномен качения (перекатывания лимфоцита вдоль сосудистой стенки). Вслед за этим устанавливается более прочное взаимодействие между р2-интегрином лимфоцитов LFA-1 и его рецептором на эндотелиальной клетке — ICAM-1. Взаимодействие усиливается благодаря «активации» р2-интегрина (т.е. изменения конформации молекулы, в результате которого повышается его сродство к рецептору — см. раздел 2.3.1.2). Это обусловливает прочное прилипание лимфоцита к сосудистой стенке. Затем под влиянием хемотаксических сигналов, поступающих из ткани
лимфоидного органа, лимфоцит начинает мигрировать из сосуда между эндотелиальными клетками.
Вышедшие из сосуда клетки попадают в примыкающие к посткапиллярным венулам тимусзависимые зоны, стромальные и дендритные клетки которых секретируют хемокины CCL19 (ELC) и С^21 (SLC), распознаваемые рецептором CCR7 на лимфоцитах. До этого момента Т- и В-клетки перемещаются в одном и том же направлении. После того как лимфоцит попадает во внутреннюю среду органа, их пути расходятся. Т-клетки остаются в тимусзависимой зоне (в паракортексе лимфатических узлов, параартериальной муфте белой пульпы селезенки и Т-зонах пейеровых бляшек). В-клетки реагируют на хемокин CXCR13 (BLC), к которому они экспрессирут рецептор СХСR5. Этот хемокин секретируют стромальные клетки фолликулов, что и определяет направление миграции В-лимфоцитов. В-лимфоциты, не несущие СХСR5, не поступают в фолликулы и локализуются в прилежащей к фолликулу зоне. Механизмы, определяющие дальнейшее перемещение лимфоцитов из органа в эфферентную лимфу, пока не установлены.
В процессе рециркуляции отсутствует «привязка» хоминга к конкретному лимфатическому узлу или пейеровой бляшке. Так, лимфоцит, покинувший шейный лимфоузел, может затем проникнуть, например, в паховый лимфатический узел или в пейерову бляшку. Принципиально лишь то, что клетки не выходят из системы вторичных лимфоидных органов. Степень специфичности хоминга минимальна в селезенке, так как лимфоциты проникают в белую пульпу при излиянии крови в краевой зоне. Только последующее перемещение в Т- и В-зоны осуществляется по описанному выше механизму.
Все сказанное выше относится к наивным лимфоцитам. После активации лимфоцитов антигеном экспрессия молекул адгезии и хемокиновых рецепторов на их поверхности существенно изменяется. Это определяет иные пути миграции клеток после контакта с антигеном. Активированные лимфоциты и клетки памяти очень слабо экспрессируют L-селектин, что обусловливает низкую вероятность их попадания в лимфатические узлы. В то же время они приобретают способность мигрировать в очаги воспаления, расположенные в нелимфоидных органах, а также в барьерные ткани (рис. 3.75). Пути миграции активированных лимфоцитов и клеток
Рис. 3.75. Мембранный фенотип Т-клеток определяет пути их миграции. Указаны молекулы адгезии, определяющие различные направления миграции Т-клеток
памяти будут специально рассмотрены далее (см. раздел 3.6.5.4). Благодаря различной экспрессии рецепторов хоминга в разных тканях формируется несколько относительно автономных путей рециркуляции. Пути рециркуляции наивных лимфоцитов и клеток памяти могут пересекаться. Так, и наивные клетки, и клетки памяти, активированные в кишечнике, попадают в брыжеечные лимфатические узлы.
Интенсивность рециркуляции Т-лимфоцтов существенно выше, чем В-лимфоцитов. У человека через каждый лимфатический узел за сутки проходит 0,3х10п, через селезенку — 2,5х10п, а через кровь — 5х10п лимфоцитов. Длительность пребывания Т-клеток в кровотоке за 1 цикл рециркуляции — около 30 мин. Благодаря рециркуляции происходит постоянное «перемешивание» лимфоидных клеток с сохранением описанного выше порядка: наивные клетки, как правило, не покидают лимфоидной ткани, тогда как клетки памяти, наоборот, часто выходят за ее пределы, мигрируя, например, в эпителиальные органы и ткани, а также в очаги воспаления. Внутри лимфоидных органов клетки также занимают соответствующие места. Упорядоченность локализации лимфоцитов различных типов очень важна для развития иммунного ответа.
Источник
Звенья патогенеза злокачественных лимфом кожи.Центральное место в патогенезе злокачественных лимфом кожи принадлежит развитию иммунопатологических процессов в коже, главным из которых является неконтролируемая пролиферация кленовых лимфоцитов. Понимание этих процессов напрямую связано с представлениями о мор-фофункциональных особенностях лимфоидной ткани кожи и ее взаимоотношениях с другими иммунокомпетентными структурами кожи в норме. В отличие от главных лимфоидных органов иммунной системы организма (тимус, селезенка, лимфатические узлы) и ряда периферических органов этой системы (лимфоглоточное кольцо носоглотки, пейеровы бляшки кишечника, легочная ткань), где имеются структурно организованные лимфоидные очаги, в коже подобным образом структурированная лимфоидная ткань отсутствует. Наличие небольших скоплений лимфоидной ткани обнаруживают лишь на местах перехода кожи к слизистым оболочкам: в наружном слуховом проходе, околососковой зоне молочных желез, генитальных областях у плодов, новорожденных и взрослых. Установлено, что лимфоидная ткань кожи осуществляет свои иммунологические функции не как фиксированная ткань, а с помощью популяции лимфоцитов, постоянно ре-циркулирующих между кожей и кровью. В эксперименте показана высокая интенсивность этой рециркуляции, которая осуществляется со скоростью 101! клеток/ч/г. Методом иммунофенотипирования с моноклональными антителами на этих лимфоцитах у здоровых лиц обнаруживают высокоаффинные, органоспецифические по отношению к коже рецепторы лимфоцитов, определяющие их аффинность к кожной ткани—CD45 RO (фенотип клеток памяти). Эти клетки обладают способностью возвращаться в те ткани, где они были первоначально стимулированы. В научной литературе для обозначения таких лимфоцитов применяют также термин «homing lymphocytes» («домашние лимфоциты»), а феномен рециркуляции лимфоцитов через кожу обозначают как «хоминговую миграцию Т-лимфоцитов в кожу». Лимфоциты, аффинные к коже, а также регионарные лимфатические узлы, куда происходит постоянная миграция из кожи «антиген-обозначенных» клеток, по аналогии с лимфоцитами, имеющими аффинитет к эпителию кишечника и легочной ткани, в настоящее время принято обозначать как «лимфоидную ткань, ассоциированную с кожей» (SALT-система). При этом некоторые авторы, учитывая топографические и функциональные особенности Т- и В-лимфоцитов в коже, различают Т-клеточную и В-клеточную лимфоидную ткань кожи. Около 16% рециркулирующих лимфоцитов, аффинных к коже, экспрессирует кожный лимфоцитарный функциональный антиген (LFA), который, как установлено с помощью молекулярной идентификации, является изофор-мой Р-селектина гликопротеина. Этот антиген определяется на лимфоцитах, мигрирующих в кожу при развитии воспалительных дерматозов (аллергический контактный дерматит, красный плоский лишай), но не определяется в лимфоцитарных инфильтратах в других тканях. У больных ЗЛК, особенно на ранних стадиях заболевания, также обнаруживают экспрессию лимфоцитами данного антигена. Однако при быстрой опухолевой прогрессии происходит потеря лимфоцитами LFA который в значительной мере определяет межклеточные взаимодействия между лимфоцитами и эпидермальными клетками, а также между лимфоцитами и эндотелиальными клетками сосудов, что приводит к ускользанию этих клеток от иммунного противоопухолевого надзора.
Феномен «хоминга» лимфоцитов кожи связан также с экспрессией ими белка, который маркируется как CD44. Благодаря этим рецепторам осуществляется связь лимфоцитов с высоким эндотелием венул поверхностного сосудистого сплетения. Повышенная экспрессия маркеров CD45 RO и CD44 определяется уже на ранних стадиях развития ЗЛК, когда процесс носит преимущественно воспалительный характер, и сохраняется по мере прогрессирования заболевания. При сравнении экспрессии маркеров CD45 RO на лимфоцитах больных различными клиническими формами ЗЛК установлено, что чаще всего они обнаруживаются у больных с эритродермическими вариантами течения этих заболеваний. С выраженной экспрессией указанных маркеров связывают плохой прогноз заболевания. В здоровой коже постоянно можно обнаружить присутствие определенного числа лимфоцитов, локализованных преимущественно вокруг посткапиллярных венул поверхностного сосудистого сплетения, в виде микрофолликулов по ходу лимфатических капилляров, а также незначительное количество (менее 1 % всех клеток эпидермиса) внутриэпидермальных лимфоцитов. Основной физиологической функцией внутриэпидермальных лимфоцитов, подобно интраэпителиальным лимфоцитам кишечника, является участие в иммунных процессах, направленных на распознавание и элиминацию антигенного материала. При хронической стимуляции лимфоцитов проонкогенными факторами уже на ранних этапах развития злокачественных лимфом кожи усиливается миграция лимфоцитов в кожу через посткапиллярные ве-нулы поверхностного сосудистого сплетения в дерме. Преимущественная локализация лимфоцитов в этих областях не случайна. В норме именно в посткапиллярных венулах наблюдается самый медленный кровоток, и, по-видимому, не случайно здесь находится зона взаимодействия лимфоцитов и эндотелия сосудов. Специализированные эндотелиальные клетки, выстилающие посткапиллярные венулы, — это органоспецифические зоны, которые благодаря комплементарным рецепторам на клеточных мембранах, распознаются лимфоцитами, обладающими аффинными к эндотелию сосудов рецепторами. У больных злокачественных лимфом кожи уже на ранних стадиях заболевания при морфологическом исследовании биоптатов кожи отмечаются пролиферация сосудов типа посткапиллярных венул, увеличение их проницаемости и формирование вокруг них инфильтратов из атипичных лимфоцитов. В дальнейшем при развитии лимфопролиферативного процесса резко усиливается селективная инфильтрация злокачественными лимфоцитами указанных зон. Непосредственный контакт между лимфоцитами и клетками специализированного высокого эндотелия посткапиллярных венул происходит с помощью адгезивных молекул эндотелиально-лимфоцитарного прилипания (ELAM-1), среди которых выделяют два важных селектина: L-селектин, который эксп-рессируется на популяции Т-лимфоцитов и представлен хоминговыми молекулами, обеспечивающими попадание Т-клеток в периферические лимфатические узлы, а также Е-селектин, который экспрессируется эндо-телиальными клетками сосудов при развитии воспалительной реакции кожи. Экспрессия этих молекул на эндотелиальных и лим-фоцитарных клетках при развитии ЗЛК возрастает, что непосредственно связано с воздействием на них цитокинов. Повышенная экспрессия адгезивных молекул эндотелиальными клетками в значительной мере определяет ин-вазивные свойства опухоли, способность к миграции опухолевых клеток в кровяное русло и, как следствие этого, плохой прогноз заболевания. Кроме экспрессии повышенного числа молекул прилипания проникновению лимфоцитов через сосуды кожи способствует также экспрессия эндотелиальными клетками иммунноассоциативных антигенов HLA-DR. Эти маркеры также способствуют обеспечению возможности лимфоцитов, преимущественно Т-клеток памяти, рециркулировать из крови в кожу и обратно и осуществлять иммунные межклеточные взаимодействия. Иммуноморфологическими методами установлено, что у здоровых людей в коже Т-лимфоциты составляют 90% всей популяции лимфоцитов. По сути дела, в настоящее время кожа рассматривается как периферический орган иммунной системы организма, в частности как зона представительства Т-лимфоцитов, наряду с паракортикальными зонами лимфатических узлов и периартериолярными зонами селезенки. В коже Т-лимфоциты сосредоточены преимущественно в эпидермисе и верхних слоях дермы. В-лимфоциты присутствуют в коже в физиологических условиях в значительно меньшем количестве и располагаются в средних и глубоких слоях дермы. В последние годы благодаря развитию иммунологии появилась возможность идентификации лимфоцитов на клеточном, молекулярном и генетическом уровне. В морфологической практике стал широко применяться метод иммунофенотипирования клеток, основанный на выявлении макромолекул, которые эксп-рессируются иммунокомпетентными клетками в процессе их дифференцировки. Эти макромолекулы получили название «CD-антитела» (от англ. clusters of differentiation — кластеры дифференцировки). С помощью моноклональных антител проводится идентификация антигенов, подтверждающих принадлежность лимфоцитов к определенной популяции, субпопуляции, а также характеризующих уровень дифференцировки лимфоцита и маркеры злокачественного роста. В таблице 1.1 представлены основные клеточные маркеры лимфоцитов по системе CD, используемые для иммунофенотипирования опухолей кожи лимфопролиферативного характера. – Также рекомендуем “Лимфоцитарная пролиферация злокачественных лимфом кожи.” Оглавление темы “Злокачественные лимфомы кожи.”: |
Источник