Характеристика рецепторов в лимфоцитов

Распознавание чужеродных агентов компонентами врожденных механизмов резистентности происходит за счет рецепторов шаблонного распознавания, которые специфичны к наиболее типичным микробным структурам, а также рецепторов к опсонинам — молекулам, «приспосабливающими относительно бедный репертуар рецепторов врожденной резистентности к антигенному разнообразию возбудителей. В связи с существенным ограничением рецепторного ресурса врожденные механизмы резистентности однотипно реагируют на разные патогены даже если последние существенно отличаются между собой. Так, например, защитные механизмы против Е. coli и S. typhi, несмотря на существенные различия между этими микроорганизмами, в целом подобны. Наиболее адекватная защита возможна только после распознавания всех антигенных детерминант, которые содержит возбудитель, что позволяет определить оптимальную силу и направленность иммунного ответа относительно данною патогена. Такое высокоспецифическое распознавание осуществляют рецепторы лимфоцитов.
Рекомбинация генетического материала
Процесс формирования рецепторов антигенного распознавания лимфоцитов является проявлением комбинационной изменчивости. При этом нуклеотидные последовательности не формируются de novo, а происходит перегруппировка уже имеющихся фрагментов ДНК. Это явление получило название рекомбинации генетического материала, или реаранжировки генов. Правда, к комбинационным механизмам приобщаются процессы, подобные мутационным изменениям. Речь идет о случайных терминальных последовательностях, которые образуют два Р-фрагмента, и встроенных нуклеотидах, формирующих N-участок. Поэтому новообразованный V-ген не является простой суммой составляющих сегментов.
К собственно механизмам комбинаторики относятся:
- Случайный выбор одной гомологической хромосомы из пары для рекомбинации сегментов V-гена.
- Случайный выбор одного сегмента из каждой группы (V, D, J).
- Случайный выбор рамки считывания участка D-сегментов; установлена возможность по крайней мере трех рамок считывания такого участка, причем в каждом случае полностью изменяется содержание всех сегментов группы (особенность триплетного кода).
- Случайное объединение субъединиц антигенраспознающего рецептора (тяжелой 31 легкой цепей В-лимфоцитов или α- и β-цепей Т-клеток), для любого из которых выполняются первые три механизма комбинаторики.
Следует отметить, что в организме существует много В-лимфоцитов с идентичными тяжелыми целями и Т-клеток с одинаковыми β-цепями. Они являются результатом пролиферации незрелых лимфоцитов с удачными «пробными» рецепторами антигенного распознавания. Тем не менее в иммунной системе каждый лимфоцит обеспечен уникальным антигенраспознающим рецептором, поскольку для его образования нужен другой компонент (соответственно — легкая цепь и α-цепь), который также сформирован случайно. Объединение двух частей рецептора создает комплекс уникальной специфичности.
За счет таких процессов организм приобретает возможность продуцировать огромное количество рецепторов антигенного распознавания, а значит — способен распознать практически любой существующий в природе антиген. Поскольку антигенсвязывающие сайты сгенерированы наугад, они не предназначены для распознавания какого-либо конкретного возбудителя. Более того, большинство из сформированных рецепторов организм никогда не использует, поскольку на протяжении существования к нему не поступают соответствующие по структуре антигены. Тем не менее большое разнообразие антигенраспознающих рецепторов обеспечивает готовность макроорганизма практически к любой антигенной агрессии, что, без сомнения, существенно повышает его шансы на выживание, поскольку поступление патогенов также является до определенной степени случайным явлением.
Рецепторы антигенного распознавания лимфоцитов состоят из двух главных частей — константной (неизменной) и вариабельной, которая является специфической к определенному антигену. Последняя уникальна у каждой клетки. Рекомбинацию претерпевают именно те гены, которые кодируют вариабельные участки рецепторов. Последние формируют антиген связывающие сайты, взаимодействующие непосредственно с эпитопами патогена.
см. Гуморальный иммунитет#Рецепторы В-лимфоцитов
Строение антител
Каркасная зона антител
Несмотря на то, что антитела различаются между собой именно по структуре вариабельного фрагмента, большая часть этого участка идентична у всех иммуноглобулинов. Речь идет о каркасной зоне. Дело в том, что все сегменты V-гена, которые входят в состав одной группы (V, D или J), содержат одинаковые последовательности нуклеотидов, которые и определяют их групповую принадлежность. При трансляции стабильных групповых последовательностей V-гена и формируется каркасная зона вариабельного участка иммуноглобулина. Материал с сайта https://wiki-med.com
Гипервариабельные участки антител
Вместе с тем каждый сегмент группы содержит специфические последовательности нуклеотидов, которые определяют его уникальность. Такие последовательности и обеспечивают формирование изменчивых участков. Поскольку весь фрагмент называется вариабельным, эти участки получили название гипервариабельных (CDR). Всего их 3 — столько же, сколько и составляющих компонентов V-гена. Поэтому каркас делится гипервариабельными участками на 4 фрагмента. Каркасная зона и гипервариабельные образования имеют разное биологическое значение. Именно каркасная часть обеспечивает формирование гидрофильного канала, который находится между вариабельными участками тяжелой и легкой цепей антитела. Образованный канал работает относительно антигенных детерминант патогена подобно капкану и является одинаковым у всех иммуноглобулинов, то есть все антитела имеют общий принцип взаимодействия с патогеном. В этом и состоит биологическое значение каркасных зон. Специфичность связывания антитела (тип распознаваемого антигена) определяется гипервариабельными участками. Поскольку они сформированы случайным образом, каждая В-клетка обеспечена антигенраcпознающим рецептором уникальной специфичности.
Рецепторы Т-лимфоцитов
см. Клеточный иммунитет#Рецепторы Т-лимфоцитов
На этой странице материал по темам:
вариабельный рецептор лимфоцитов
строение и роль рецепторов атиген- распознающих рецепторов т лимфоцитов
рецепторы в-лимфоцитов
рецепторы для т-лимфоцитов это
атлас схема строения рецепторного аппарата т лимфоцитов
Источник
Оглавление темы “Иммунная система. Функции клеток иммунной системы. Т-лимфоциты. В-лимфоциты. Структура и функции органов иммунной системы.”:
1. Иммунная система. Иммунная система человека. Неспецифический иммунитет. Специфический иммунный ответ. Рециркуляция лимфоцитов.
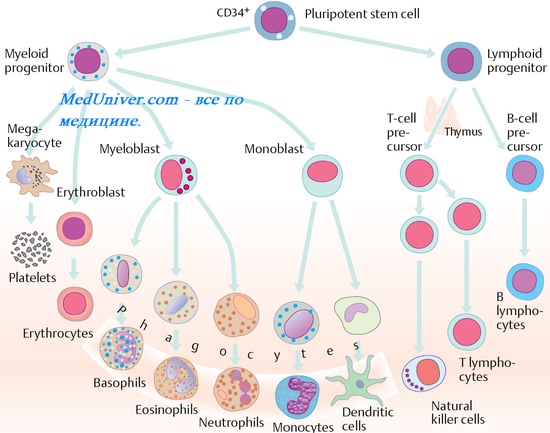
2. Происхождение ( образование ) клеток иммунной системы. Функции клеток иммунной системы. Лимфопоэз. Бурса Фабрициуса.
3. Т-лимфоциты. Характеристика Т-лимфоцитов. Типы молекул на поверхности Т-лимфоцитов.
4. Популяции Т-лимфоцитов. Субпопуляции Т-лимфоцитов. СD4 Т-лимфоциты. СD8 Т-лимфоциты.
5. Функции Т-лимфоцитов. Активированные Т-лимфоциты. Цитокины.
6. В-лимфоциты. Характеристика В-лимфоцитов. Клетки памяти.
7. Функции В-лимфоцитов. Типы молекул на поверхности В-лимфоцитов.
8. Антигенпредставляющие клетки. Структура и функции органов иммунной системы.
9. Костный мозг. Функции костного мозга. Миеломоноцитопоэз.
10. Тимус (вилочковая железа). Тимоциты. Функции тимуса ( вилочковой железы ). Этапы отбора тимоцитов.
В-лимфоциты. Характеристика В-лимфоцитов. Клетки памяти.
Предшественники В-лимфоцитов из эмбриональной печени человека на 8—9-й неделе развития плода перемещаются в костный мозг, где они созревают под влиянием интерлейкина 7, который секретируют стромальные клетки.
В ранний период созревания в костном мозге формируется и фиксируется на поверхностной мембране В-лимфоцита иммуноглобулиновый антигенраспознающий рецептор. Незрелый В-лимфоцит уже несет мембранные В-клеточные рецепторы (IgM) определенной специфичности, т. е. способные распознать и связать конкретный антиген. При встрече незрелых В-лимфоцитов с аутоантигенами связывание последних с мембранными IgM ведет к остановке дифференцировки этих В-лимфоцитов, с последующей их гибелью путем апоптоза. На этой стадии погибают до 90 % незрелых В-клеток. Часть незрелых В-клеток после контакта с растворимым аутоанти-геном не погибают, а переходят в состояние анергии по отношению к данному антигену. Этим объясняется врожденное состояние иммунологической толерантности (неспособности ответить) на собственные аутоантигены.
Выжившие в этом процессе отбора В-клетки по мере созревания покидают костный мозг, переходя в кровяное русло через венозные синусы, и поступают в периферические органы иммунной системы, где превращаются в зрелые В-лимфоциты. В кровяном русле В-лимфоциты включаются в рециркуляцию и заселяют В-зоны периферических органов и тканей иммунной системы.
Около 2 • 107 В-лимфоцитов покидают костный мозг ежедневно, что составляет 5—10 % общей популяции В-клеток периферической крови. Этого достаточно, чтобы полностью обновить популяцию периферических В-лимфоцитов за 4—5 дней. Среди сохранившихся и вступивших в рециркуляцию относительно долгоживущих В-лимфоцитов (с полупериодом жизни 1—2 мес) значительную часть составляют клетки памяти, способные быстро ответить пролиферацией, активацией, продукцией и секрецией специфических антител (иммуноглобулинов) на повторную встречу с тем же антигеном.
– Также рекомендуем “Функции В-лимфоцитов. Типы молекул на поверхности В-лимфоцитов..”
Источник
Оглавление темы “СD8 лимфоциты. Антиген ( Аг ) представляющие клетки. Классификация антигенов ( Аг ).”:
1. Рецептор Т-клеток. Активация Т-лимфоцитов. Субпопуляции Т-лимфоцитов. СD4+ лимфоциты.
2. СD8 лимфоциты. Т супрессоры. Цитотоксические Т-лимфоциты ( ЦТЛ ). Т-киллеры. Цитотоксический эффект Т-киллеров. Т-клетки памяти.
3. В-лимфоциты. В-клетки. Созревание В-лимфоцитов. Продолжительность жизни B-лимфоцитов. Маркеры В-клеток ( В-лимфоцитов ).
4. Естественные киллеры ( NK-клетки ). Перфорины. Функция естественных киллеров. Задачи NK-клеток.
5. Антиген ( Аг ) представляющие клетки. Иммунные реакции. Классификация имунных реакций. Антигены. Свойства антигенов. Структура антигенов ( Аг ). Валентность Аг.
6. Классификация антигенов ( Аг ). Иммуногены. Гаптены. Непреципитирующие гаптены. Преципитирующие гаптены. Полугаптены. Проантигены.
7. Адъюванты. Адъювант Фройнда. Толерогены. Природа антигенов (Аг). Происхождение антигенов. Молекулярная масса антигенов (Аг).
8. Специфичность антигенов (Аг). Видовые антигены. Групповые антигены (Аг). Гетерогенные антигены (Аг). Антиген Форссмана. Аллоантигены ( изоантигены ).
9. Патологические антигены. Аутоантигены. Врождённые аутоантигены. Приобретённые аутоантигены. Суперантигены.
10. Антигены ( Аг ) микроорганизмов. Антигены бактерий. Капсульные антигены ( К-антигены ( Аг )). Соматические антигены ( O-антигены ( Аг )). Жгутиковые антигены ( H-антигены ( Аг )). Vi-Ar ( Антиген вирулентности ).
В-лимфоциты. В-клетки. Созревание В-лимфоцитов. Продолжительность жизни B-лимфоцитов. Маркеры В-клеток ( В-лимфоцитов ).
Известны субпопуляции В-клеток: предшественники антителообразующих (плазматических) клеток и В-клетки памяти (эффекторы вторичных иммунных реакций). Доминирующую субпопуляцию составляют предшественники антителообразующих клеток, дифференцирующиеся после антигенной стимуляции в плазматические клетки (плазмоциты), синтезирующие Ig.
Созревание В-лимфоцитов
Из костного мозга пре-В-клетки мигрируют в тимуснезависимые зоны лимфоидных органов. Так, в физиологических условиях в селезёнке В-лимфоциты располагаются в краевой зоне белой пульпы, в лимфатических узлах — в наружной зоне кортикального слоя, где они формируют зародышевые центры фолликулов. Сигналы, определяющие судьбу и дифференцировку этих иммунокомпетентных клеток, поступают из красного костного мозга, стромальных клеток и других клеток иммунной системы.
На периферии (вне костного мозга) В-лимфоциты приобретают характерные для них поверхностно-клеточные маркёры. Продолжительность жизни В-лимфоцитов различна — от многих лет (В-клетки памяти) до нескольких недель (клоны плазматических клеток).
После антигенной стимуляции В-лимфоциты дифференцируются в плазматические клетки (интенсивно синтезирующие и секретирующие AT) и В-клетки памяти. Плазматические клетки синтезируют Ig того же класса, что и мембранный Ig В-лимфоцита-предшественника.
Маркеры В-клеток ( В-лимфоцитов )
Основные маркёры В-лимфоцитов — мембранные Ig при этом клетки одного клона (быстро формирующегося в результате серии последовательных делений потом-ства одной В-клетки ) экспрессируют молекулы Ig, специфически связывающие только один эпитоп Аг. Такие клетки синтезируют моноклональные AT, способные распознавать и связывать только один Аг. Аг-связывающий участок мембранного Ig В-лимфоцита играет роль клеточного Аг-распознающего рецептора.
Помимо мембранных Ig, В-лимфоцит несёт другие маркёры; рецепторы Fc-фрагмента Ig, CD10 {на незрелых В-клетках), CD19, CD20, CD21, CD22, CD23 (вероятно, участвуют в клеточной активации), рецепторы к С3b и C3d, молекулы МНС классов I и И.
– Также рекомендуем “Естественные киллеры ( NK-клетки ). Перфорины. Функция естественных киллеров. Задачи NK-клеток.”
Источник
Выделяют несколько субпопуляций В-клеток: В1, В2 и В клетки маргинальной зоны (MZB). Основная из них — В2-лимфоциты, или «обычные» В-клетки. Практически все данные о В-лимфоцитах получены на В2-клет- ках. Им посвящен материал, представленный в двух первых разделах главы. Другие субпопуляции будут рассмотрены в специальном разделе (3.1.1.3).
Основное свойство В-лимфоцитов — экспрессия иммуноглобулинового рецептора для распознавания антигенов — BCR. На поверхности зрелой В-клетки содержится около 150 000 комплексов BCR.
Напомним, что на мембране зрелой наивной В-клетки (т.е. В-клетки, ранее не контактировавшей с антигеном), содержатся иммуноглобулины классов IgM и IgD. Если IgM-антитела, секретируемые клеткой, являются пентамерами (т.е. содержит 5 мономерных структур IgM), мембранный IgM В- клеточного рецептора представляет собой мономер. Для зрелых В2-клеток характерна низкая экспрессия мембранного IgM и высокая — IgD. После активации антигеном (т.е. в ходе иммунного ответа) класс антигенраспозна- ющего рецептора В-клетки может изменяться: вместо IgM и IgD на мембране появляются иммуноглобулины других классов — IgG, IgE и IgA. Н-цепи мембранных иммуноглобулинов отличаются от соответствующих молекул растворимых иммуноглобулинов-антител наличием на С-конце двух дополнительных участков — трансмембранного и цитоплазматического. При помощи трансмембранного участка молекула иммуноглобулина встраивается в мембрану клетки (о переключении изотипов, а также переходе от мембранной к растворимой форме иммуноглобулинов см. раздел 3.1.4.4).
Антигенраспознающий рецептор В-клеток (BCR) уже рассматривался выше (см. рис. 3.7). В состав BCR входит также ряд молекул, относящихся к суперсемейству иммуноглобулинов. С мембранными иммуноглобулинами нековалентно связаны 2 пары молекул — гетеродимеры, содержащие полипептидные цепи Iga (CD79a) и IgP ^D79b). Обе полипептидные цепи встроены в мембрану В-лимфоцита. Их цитоплазматическая часть контактирует с тирозинкиназами Fyn, Lck, Blk, что позволяет им участвовать в передаче сигнала о связывании антигена внутрь клетки. С BCR ассоциировано еще несколько мембранных молекул — CD19, CD21 (рецептор для комплемента CR2) и CD81. Они не являются интегральной частью рецептора, но при взаимодействии с антигеном между ними и В-клеточным рецептором устанавливается связь, и они вносят существенный вклад в усиление активационного сигнала, поступающего в клетку от рецептора. Особенно четко это показано для молекулы CD21, являющейся рецептором для комплемента (CR2) и связывающего фрагмент C3b после взаимодействия с антигеном (см. 3.6.2.1). Молекулы, связанные с BCR, рассматривают как маркеры В-лимфоцитов и их экспрессию определяют (с помощью моноклональных антител) для подсчета численности В-клеток. Однако строгоспецифичной для В-лимфоцитов является только молекула CD19. Маркером фолликулярных, В2-клеток является мембранная молекула CD23.
На поверхности В-лимфоцитов конститутивно или под влиянием активации экспрессируются также молекулы, необходимые для выполнения функций, не связанных с распознаванием антигена и выработкой антител. Так, В-клетки несут на поверхности молекулы MHC не только I, но и II класса, а также костимулирующие молекулы CD40, CD86, а при активации — также CD80. Благодаря экспрессии этих молекул В-лимфоциты могут выполнять роль «профессиональных» АПК. В-клетки экспрессируют молекулы адгезии (Р1-интегрины VLA-2 и VLA-4, в2-интегрин LFA-1, L-селектин CD62L и др.), позволяющие им мигрировать из сосудов и перемещаться в тканях. Присутствие на их поверхности Fc-рецепторов (FcyRI№ — CD32) и уже упомянутых рецепторов для комплемента (CR2) в регуляции активности В-клеток играет большую роль, чем для выполнения ими эффекторных функций.
В-клетки экспрессируют многочисленные рецепторы для цитокинов, из которых наиболее важны рецепторы для IL-4, IL-5, IL-6, IL-2, IL-1, IL-10 и некоторых других. На их поверхности присутствуют рецепторы для цитокинов семейства TNF: BAFF (B-cell activating factor of TNF family) — BAFF-R, BCMS, TAC-1, а также APRIL (A proliferation inducing ligand) — HSPG. Эти цитокины защищают В-клетки от развития апоптоза и выполняют гомеостатическую функцию, поддерживая численность этих клеток на постоянном уровне. На В-лимфоцитах представлены рецепторы для хемокинов: например, CXCR4 (для SDF-1), CXCR5 (для BLC, служащего основным хемоаттрактантом для наивных В-клеток), CCR3 (для эотаксинов), CCR6 (для LARC).
Главное средоточие В2-клеток — лимфоидные фолликулы — наиболее универсальная лимфоидная структура, которая может входить в состав вторичных лимфоидных органов или существовать самостоятельно. В связи с этим В2-клетки иногда называют фолликулярными В-лимфоцитами. В2-клетки выявляют в костном мозгу, в пространстве вокруг синусоидов. Кроме того, В-клетки, относящиеся к различным субпопуляциям, присутствуют в значительных количествах в межфолликулярных областях, в мозговых шнурах лимфатических узлов, краевых зонах белой пульпы селезенки. В виде диффузно распределенных клеток В-лимфоциты представлены в соединительнотканных отделах барьерных тканей — дерме, собственной пластине слизистых оболочек, подслизистом слое. В2-лимфоциты рециркулируют, хотя и значительно слабее, чем Т-клетки. Их содержание в кровотоке невелико: по данным разных авторов, В-клетки составляют 10—13% от общего числа лимфоцитов крови (нормальный разброс — 5—25%). Абсолютное содержание В-лимфоцитов в крови составляет 110—375х109/л.
Ранее В-лимфоциты считали короткоживущими клетками. Действительно, вне фолликулов В2-клетки живут около недели. Однако в их естественном микроокружении В-клетки способны существовать достаточно долго — в течение нескольких недель и даже месяцев. Срок полуобновления пула В-клеток при действии повреждающих факторов составляет 13 сут. Главные факторы, поддерживающие жизнеспособность В-лимфоцитов, — цитокины семейства TNF — BAFF и APRIL.
Источник