Где происходит созревание лимфоцитов

Оглавление темы “СD8 лимфоциты. Антиген ( Аг ) представляющие клетки. Классификация антигенов ( Аг ).”:
1. Рецептор Т-клеток. Активация Т-лимфоцитов. Субпопуляции Т-лимфоцитов. СD4+ лимфоциты.
2. СD8 лимфоциты. Т супрессоры. Цитотоксические Т-лимфоциты ( ЦТЛ ). Т-киллеры. Цитотоксический эффект Т-киллеров. Т-клетки памяти.
3. В-лимфоциты. В-клетки. Созревание В-лимфоцитов. Продолжительность жизни B-лимфоцитов. Маркеры В-клеток ( В-лимфоцитов ).
4. Естественные киллеры ( NK-клетки ). Перфорины. Функция естественных киллеров. Задачи NK-клеток.
5. Антиген ( Аг ) представляющие клетки. Иммунные реакции. Классификация имунных реакций. Антигены. Свойства антигенов. Структура антигенов ( Аг ). Валентность Аг.
6. Классификация антигенов ( Аг ). Иммуногены. Гаптены. Непреципитирующие гаптены. Преципитирующие гаптены. Полугаптены. Проантигены.
7. Адъюванты. Адъювант Фройнда. Толерогены. Природа антигенов (Аг). Происхождение антигенов. Молекулярная масса антигенов (Аг).
8. Специфичность антигенов (Аг). Видовые антигены. Групповые антигены (Аг). Гетерогенные антигены (Аг). Антиген Форссмана. Аллоантигены ( изоантигены ).
9. Патологические антигены. Аутоантигены. Врождённые аутоантигены. Приобретённые аутоантигены. Суперантигены.
10. Антигены ( Аг ) микроорганизмов. Антигены бактерий. Капсульные антигены ( К-антигены ( Аг )). Соматические антигены ( O-антигены ( Аг )). Жгутиковые антигены ( H-антигены ( Аг )). Vi-Ar ( Антиген вирулентности ).
В-лимфоциты. В-клетки. Созревание В-лимфоцитов. Продолжительность жизни B-лимфоцитов. Маркеры В-клеток ( В-лимфоцитов ).
Известны субпопуляции В-клеток: предшественники антителообразующих (плазматических) клеток и В-клетки памяти (эффекторы вторичных иммунных реакций). Доминирующую субпопуляцию составляют предшественники антителообразующих клеток, дифференцирующиеся после антигенной стимуляции в плазматические клетки (плазмоциты), синтезирующие Ig.
Созревание В-лимфоцитов
Из костного мозга пре-В-клетки мигрируют в тимуснезависимые зоны лимфоидных органов. Так, в физиологических условиях в селезёнке В-лимфоциты располагаются в краевой зоне белой пульпы, в лимфатических узлах — в наружной зоне кортикального слоя, где они формируют зародышевые центры фолликулов. Сигналы, определяющие судьбу и дифференцировку этих иммунокомпетентных клеток, поступают из красного костного мозга, стромальных клеток и других клеток иммунной системы.
На периферии (вне костного мозга) В-лимфоциты приобретают характерные для них поверхностно-клеточные маркёры. Продолжительность жизни В-лимфоцитов различна — от многих лет (В-клетки памяти) до нескольких недель (клоны плазматических клеток).
После антигенной стимуляции В-лимфоциты дифференцируются в плазматические клетки (интенсивно синтезирующие и секретирующие AT) и В-клетки памяти. Плазматические клетки синтезируют Ig того же класса, что и мембранный Ig В-лимфоцита-предшественника.
Маркеры В-клеток ( В-лимфоцитов )
Основные маркёры В-лимфоцитов — мембранные Ig при этом клетки одного клона (быстро формирующегося в результате серии последовательных делений потом-ства одной В-клетки ) экспрессируют молекулы Ig, специфически связывающие только один эпитоп Аг. Такие клетки синтезируют моноклональные AT, способные распознавать и связывать только один Аг. Аг-связывающий участок мембранного Ig В-лимфоцита играет роль клеточного Аг-распознающего рецептора.
Помимо мембранных Ig, В-лимфоцит несёт другие маркёры; рецепторы Fc-фрагмента Ig, CD10 {на незрелых В-клетках), CD19, CD20, CD21, CD22, CD23 (вероятно, участвуют в клеточной активации), рецепторы к С3b и C3d, молекулы МНС классов I и И.
– Также рекомендуем “Естественные киллеры ( NK-клетки ). Перфорины. Функция естественных киллеров. Задачи NK-клеток.”
Источник
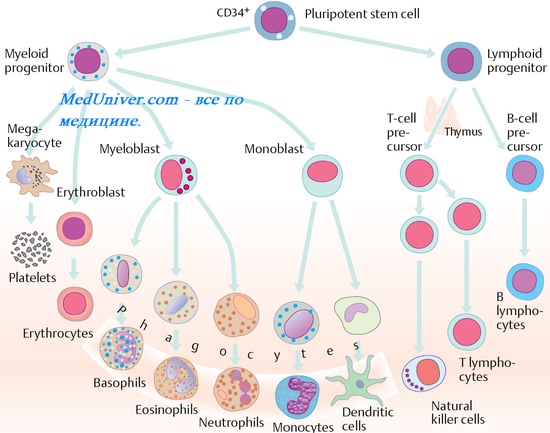
Часть лимфоидных стволовых клеток мигрирует в вилочковую
железу (тимус) и созревает там до Т-лимфоцитов; другая их часть остается в
костном мозге и созревает до В-лимфоцитов. На пути созревания (дифференцировки)
в центральных органах иммунитета стволовая клетка проходит несколько этапов.
Эти этапы созревания Т- и В-лимфоцитов происходят без участия антигена и
поэтому получили название антиген независимой дифференцировки.
Первый этап – это появление на стволовой клетке структур, указывающих,
по какому пути дифференцировки (Т- или В-лимфоцитов) пойдет ее развитие; он
происходит еще в костном мозге.
Ранний предшественник В-лимфоцитов имеет на своей мембране так
называемую суррогатную L-цепь молекулы иммуноглобулина, а ранний предшественник
Т-лимфоцитов – гликопротеин с молекулярной массой 3,3*104 D (ГП-33), который
впоследствии ассоциируется с бета-цепью Т-клеточного антигенораспознающего
рецептора.
Указанные структуры, определяющие дальнейшую судьбу
гемопоэтической стволовой клетки, играют важную роль: 1) задают путь
дифференцировки Т- и В-лимфоцитов; 2) передают сигнал ранним предшественникам
Т- и В-лимфоцитов, по которому начинается их пролиферация.
Например, на этапе появления суррогатной L-цепи и ГП-33
таких клеток в костном мозге и тимусе у мышей насчитывается по 3-5*106. Однако
уже через 4 недели их количество увеличивается в 10-100 раз и составляет 5*107
– 108 клеток. Таким образом, наличие суррогатной L-цепи и ГП-33 является тем
инструментом, с помощью которого количество пре-В- и пре-Т-клеток достигает
необходимого уровня.
Второй этап дифференцировки лимфоцитов- это появление
незрелых предшественников В- и Т-лимфоцитов.
Он определяется появлением на мембране В- и Т-лимфоцитов
антигенраспознающих рецепторов, с помощью которых В- и Т-лимфоциты после этого
этапа дифференцировки приобретают способность распознавать антигены.
Для В-лимфоцита антигенраспознающим рецептором (ВАГРР)
является мембранный IgM, а для Т-лимфоцита (ТАГРР) – специальная димерная молекула,
имеющая альфа- и-бетацепи, и относящаяся к супер семейству иммуноглобулинов
(подробно об антигенраспознающих рецеп-торах будет сказано в разделах о Т- и
В-лимфоцитах).
Между I и II этапом есть еще несколько стадий дифференцировки,
о которых здесь не упоминается.Таким образом, появление на поверхности ранних
предшественников лимфоидных клеток определенных структур (рецепторов) служит
тем сигналом, который позволяет клеткам дифференцироваться в специфическую
линию лимфоцитов. Имеющие такие рецепторы клетки-предшественники мигрируют в
определенную область центральных иммунных органов, а затем взаимодействуют со
специфическим микроокружением, необходимым для того, чтобы в этой области
произошла дифференцировка данной клетки. В ответ на контакт с
клеткой-предшественником, в стромальных клетках локального микроокружения в
свою очередь развиваются молекулярные процессы, которые направлены на
“обучение” (коммитирование) клеток-предшественников для дальнейшей их
дифференцировки в отдельно взятую линию.
Показано, что клетки-предшественники в тесном контакте с их
стромальным клеточным ми-кроокружением сохраняют очередную стадию
дифференцировки до тех пор, пока имеется достаточно пространства в соответствующей зоне (компартменте)
центрального органа иммунитета (тимуса или костного мозга), и пока
соответствующий цитокин, ответственный за пролифера-цию и дифференцировку
продуцируется в данной зоне.
Таким образом, отсутствие контакта со стромальными клетками
и потеря сигнала через ци-токиновый рецептор являются теми условиями, при
которых клетка-предшественник переходит к дальнейшей стадии дифференцировки.
Это позволяет ей осуществить новый контакт в следующем, новом месте
микроокружения.
Источник
Т- и В-лимфоциты, которые покидают соответственно вилочковую железу и костный мозг, находятся на ранней стадии иммунологического созревания. Когда антигенпрезентирующие клетки представляют антиген этим Т- и В-клеткам в первый раз, например, в лимфатическом узле или селезенке, лимфоциты трансформируются в Т- или В-иммунобласты.
Иммунобласты являются самыми крупными лимфоидными клетками. Они имеют светлое ядро с тонкоструктурированным хроматином и крупными одиночными ядрышками. Цитоплазма иммунобластов выглядит как широкий базофильный ободок. Т-иммунобласты либо выполняют свою эффекторную Т-клеточную функцию и погибают, либо становятся Т-клетками памяти. Клетки последнего типа более сильно и интенсивно отвечают на стимуляцию при контакте с тем же самым антигеном.
В-иммунобласты превращаются в плазматические клетки, проходя перед этим стадию плазмоцитоидных клеток. Первоначально плазмоцитоидные клетки имеют морфологические признаки, промежуточные между малыми лимфоцитами и зрелыми плазматическими клетками, на этой стадии они секретируют преимущественно IgM. При развитии размер клеток увеличивается, и они приобретают черты типичных плазматических клеток, продуцирующих иммуноглобулины различных классов.
Плазматические клетки крупнее, чем лимфоциты. Они имеют эксцентрично расположенное круглое ядро. Цитоплазма плазматических клеток строго базофильна, за исключением светлой перинуклеарной зоны, занятой комплексом Гольджи. При первичном иммунном ответе плазмоцито-идные лимфоциты преобладают, а ассоциированная с ними продукция иммуноглобулинов невелика.
Пути созревания В-лимфоцитов в лимфатическом узле после выхода из костного мозга
В лимфатических узлах и других лимфоидных органах, прежде всего селезенке, в ответ на антигенную стимуляцию Т-зависимыми антигенами образуются зародышевые центры. Первоначально В-клетки, из которых формируется зародышевый центр, получают активирующий сигнал вне фолликулов, в Т-клеточных зонах от интердигитальных дендритных клеток и Т-хелперов. Каждый фолликул колонизируют в среднем три В-клеточных бласта. Эти бласты подвергаются массивной клональной экспансии, и в них активизируется механизм соматических гипермутаций, который действует на вариабельные области генов иммуноглобулинов.
Зрелые зародышевые центры разделены на темную и светлую зоны. Темную зону занимают пролиферирующие бласты, называемые центробластами, затем из них формируются центроциты, которые не находятся в митотическом цикле и вытесняются в светлую зону. Два типа этих лимфоидных клеток известны как клетки фолликулярных центров. Центробласты — крупные клетки, но обычно мельче иммунобластов.
Светлая зона содержит сеть фолликулярных дендритных клеток, которые имеют способность поглощать и процессировать (обрабатывать) антиген. Антигенные детерминанты могут удерживаться на поверхности дендритных клеток более года или в форме иммунного комплекса, или в нативном необработанном виде. Антиген может также поглощаться В-клетками, которые могут процессировать его и представлять Т-клеткам.
Центроциты селектируются по их способности взаимодействовать с антигеном, удерживаемым дендритными клетками. Для центроцитов характерна высокая смертность путем апоптоза. Перекрестное связывание иммуноглобулиновых рецепторов со специфическим антигеном при одновременном проведении сигнала через рецептор CD40 отменяет апоптоз центроцитов. После получения сигналов через перечисленные рецепторы на центроцитах увеличивается экспрессия поверхностных иммуноглобулинов, и они приобретают характеристики В-клеток памяти.
Зародышевые центры сохраняются около трех недель после иммунизации, а после этого В-бласты памяти продолжают пролиферировать в фолликулах в течение месяцев, пока длится Т-зависимый антительный ответ. Эти клетки, вероятно, являются источником и плазматических клеток, и клеток памяти, необходимых для поддержания долгосрочной продукции антител и иммунологической памяти после первых трех недель Т-зависимого антительного ответа.
Описанные на нашем сайте реактивные лимфоидные клетки встречаются главным образом в лимфоидных тканях, их также можно увидеть в крови при инфекционном мононуклеозе и других вирусных инфекциях, а также у больных неходжкинскими лимфомами (в случае лейкемизации).
– Также рекомендуем “Гематология: Рециркуляция лимфоцитов”
Оглавление темы “Гематология”:
- Гематология: История изучения крови
- Концепция единства кровяной ткани. Кровь как мезенхимальная ткань
- Гематология: Эмбриология крови – кровяной ткани
- Гематология: Клетки и цитокины регулирующие развитие В-клеток крови
- Гематология: Развитие Т-лимфоцитов и его регуляция
- Гематология: Развитие лимфоцитов в лимфатических узлах и селезенке
- Гематология: Рециркуляция лимфоцитов
- Клиническое обследование пациента в гематологии: сбор анамнеза, объективный осмотр
- Морфологическое исследование клеток крови. Причины изменений
- Гематология: Лейкоцитарная формула в норме и при болезнях
Источник
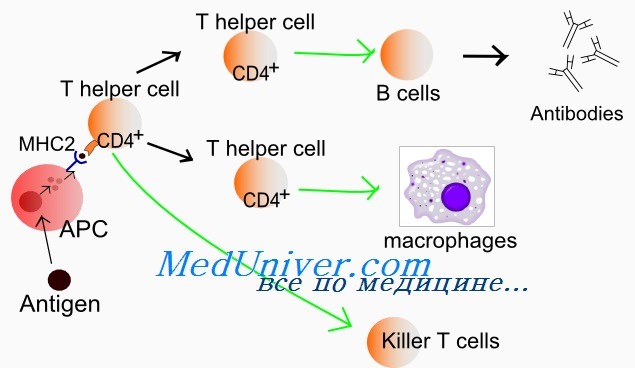
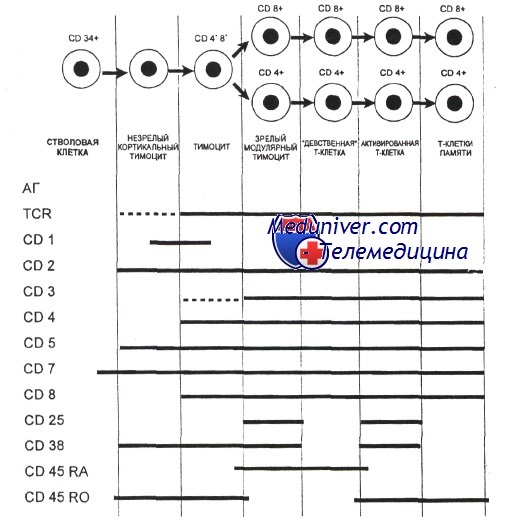
Гематология: Развитие Т-лимфоцитовПредшественники, прекоммитированные к развитию в Т-лимфоциты, непрерывно мигрируют от костного мозга в вилочковую железу, однако их число, по-видимому, очень мало. Незрелые предшественники Т-клеток поступают в область коры вилочковой железы, где созревают в функциональные субпопуляции Т-клеток. Особенностью развития тимоцитов является высокая скорость пролиферации. Различные стадии развития тимоцита можно определить по изменению экспрессии поверхностных и цитоплазматических молекул, рецепторов цитокинов и состояния реаранжировки генов Т-клеточного рецептора. Т-клеточный рецептор (TCR) распознает только короткие пептиды, которые заполняют полость в молекулах первого или второго класса главного комплекса гистосовместимости (МНС I и II, МНС комплекс у человека обозначается как HLA). Презентация таких пептидов Т-лимфоцитам обеспечивается антигенпредставляющими клетками. TCR — гетеродимерный комплекс, сформированный при ассоциации а-и бета- или у- и S-субъединиц. Каждая из этих субъединиц кодируется отдельным геном. Т-клетки с рецепторами типов ар и уq являются различными линиями, которые разделяются до начала реаранжировки гена TCR. Наименее зрелые клетки-предшественники в вилочковой железе экспрессируют антиген CD7. Некоторые из них экспрессируют также CD44. CD44, рецептор для гиалуроновой кислоты, — один из факторов, определяющих перемещение предшественников Т-клеток в вилочковую железу. Из этих предшественников далее образуются CD2+/CD7+ клетки, в цитоплазме которых присутствует CD3. Белковый комплекс CD3 обеспечивает передачу сигналов через TCR. Пептидные цепи CD3-комплекса начинают синтезироваться на стадии про-Т-лимфоцита, и их экспрессия предшествует появлению на мембране TCR. Т-лимфоциты уq-линии первыми начинают экспрессировать рецептор CD3, но на них нет молекул CD4 или CD8. Несколько позднее на поверхности клеток, экспрессирующих рецептор типа аb, начинают одновременно экспрессироваться антигены CD4 и CD8. К этому времени заканчивается реаранжировка генов а- и р-цепей, и на поверхности клеток экспрессируется рецепторный комплекс TCR ab/CD3. По мере того как клетки, экспрессирующие CD4+, CD8+ и TCR (так называемые дважды положительные клетки), проходят от коры к мозговому веществу вилочковой железы, они созревают в CD4+ хелперные или CD8+ цитотоксические Т-клетки. Клетки, которые созревают до стадии CD4+ или CD8+, составляют менее 5% тимоцитов. Эти лимфоциты покидают вилочковую железу и заполняют вторичные лимфоидные ткани (лимфатические узлы, селезенку, лимфоидную ткань, ассоциированную со слизистыми оболочками). Т-лимфоциты с рецептором уq и CD5+B1-клетки являются функционально аналогичными популяциями, которые развиваются параллельно. Т-клетки с рецептором типа уq найдены в различных тканях, включая селезенку, эпидермис и эпителий слизистой оболочки матки, влагалища и языка. Предполагается, что эта популяция клеток может исполнять роль иммунного надзора в перечисленных тканях.
Формирование комплекса Т-клеточного рецептора и положительная и отрицательная селекция в вилочковой железеКак и гены иммуноглобулинов, гены Т-клеточного рецептора собираются из различных сегментов, находящихся первоначально в зародышевой конфигурации. Гены Т-клеточного рецептора подвергаются процессу соматической реаранжировки, при которой кодирующие сегменты присоединяются друг к другу, а присутствующие между ними интронные последовательности удаляются. Вариабельность соединений и вставок обеспечивает возникновение различий TCR и приводит к огромному количеству различных последовательностей гипервариабельного участка TCR. Репертуар различных TCR считается даже большим, чем репертуар молекул Ig (по оценкам, репертуар ab-TCR составляет 1015, а для yq-TCR — 1018). В отличие от В-лимфоцитов, Т-лимфоциты не секретируют свои рецепторы. Дважды позитивный тимоцит проходит через многоступенчатый процесс, известный как тимическая селекция. На первой фазе, называемой положительной селекцией, TCR дважды позитивного тимоцита взаимодействуют с молекулами МНС, которые экспрессируются эпителиальными клетками в коре вилочковой железы. Тимоциты, способные распознавать комплекс антигена с молекулой HLA, ускользают от апоптоза и вступают в дальнейшую дифференцировку, в то время как тимоциты, не способные к такому взаимодействию, погибают. Поскольку рекомбинации, затрагивающие возникновение TCR, являются случайными, в вилочковой железе могут развиваться и пройти позитивную селекцию Т-клетки, экспрессирующие TCR, специфичные к чужеродным и собственным антигенам. Существует вероятность, что Т-клетки, реактивные к собственным структурам организма, будут взаимодействовать с аутоантигенами в тканях, что может привести к нежелательным аутоиммунным реакциям. Для предотвращения этого дважды положительные клетки подвергаются второй фазе отбора, которая называется негативной селекцией. В ходе этой фазы дважды положительные клетки взаимодействуют с комплексом МНС-пептид, экспрессированным на поверхности дендритной клетки. Т-клетки, TCR которых взаимодействует с комплексом МНС-пептид с высокой аффинностью, подвергаются апоптозу. В ходе негативной селекции удаляются Т-клетки, экспрессирующие TCR, обладающие реактивностью к собственным антигенам. Дважды положительные клетки, пережившие негативную селекцию, снижают экспрессию корецепторов CD4 или CD8, что приводит к развитию или CD4+CD8″ или CD4″CD8+ (моноположительных) Т-лимфоцитов. При этом Т-лимфоциты, распознающие антигены, представляемые молекулами МНС II класса, сохраняют экспрессию корецептора CD4+, a Т-лимфоциты, распознающие антигены, представляемые молекулами МНС I класса, сохраняют экспрессию корецептора CD8+. Эти клетки покидают вилочковую железу и образуют периферические зрелые CD4+ и CD8+ Т-клетки.
Клетки и цитокины, регулирующие развитие Т-клетокВилочковая железа состоит из корковой и медуллярной областей, различающихся по количеству тимоцитов и составу стромальных клеток. Стромальные клетки вилочковой железы влияют на развитие тимоцитов как посредством прямых межклеточных взаимодействий, так и секрецией растворимых медиаторов. Тимоциты и клетки тимического эпителия экспрессируют ряд поверхностных клеточных детерминант, некоторые из которых вовлечены в адгезию этих клеточных популяций друг к другу. Рецептор CD2 на тимоцитах определяет связывание с детерминантой CD58 (LFA-3) и межклеточной адгезионной молекулой-1 (ICAM-1), представленными на клетках тимического эпителия. Взаимодействия между развивающимися лимфоцитами и стромой могут активировать обе популяции. Например, связывание тимоцитов со стромой может стимулировать продукцию ИЛ-1 стромальными клетками и повышать экспрессию рецептора ИЛ-2 на тимоцитах. ИЛ-7 стимулирует полиферацию тимоцитов, и фактор стволовых клеток увеличивает этот эффект. Эпителиальные клетки вилочковой железы у человека являются источником цитокинов ИЛ- 1а и b, ИЛ-3, ИЛ-6, ИЛ-8, колониестимулирующих факторов (Г-КСФ и ГМ-КСФ), лейкозингибирующего фактора (LIF) и TGF-p, а также гормонов тимозина или тимопоэтина, оказывающих влияние на пролиферацию и дифференцировку тимоцитов. Цитокины, необходимые для развития Т-клеток, могут продуцироваться самими Т-клетками. Тимоциты производят IFN-y, ФНО-а, ИЛ-2, ИЛ-3 и ИЛ-4. Естественные киллерные клетки. Естественные киллеры (NK) обладают способностью спонтанно лизировать некоторые опухолевые клетки-мишени. Человеческие NK-клетки экспрессируют CD16 и CD56, но не имеют TCR или CD3. По морфологическим характеристикам эти клетки представляют собой большие гранулярные лимфоциты (в них присутствуют крупные цитоплазматические гранулы). NK-клетки имеют гемопоэтическое происхождение и развиваются в костном мозге, но пока мало известно об их предшественниках. – Также рекомендуем “Гематология: Развитие лимфоцитов в лимфатических узлах и селезенке” Оглавление темы “Гематология”:
|
Источник