Дифференцировка т и в лимфоцитов презентация

1. Дифференцировка Т-лимфоцитов
Выполнила
Казьмина Н.А
3.4.12б
2.
3.
Относительное содержание Т-лимфоцитов в крови составляет
в среднем около 73% (55-85%) от общего числа лимфоцитов;
абсолютное — (950—2100)х109 клеток в литре;
На поверхности αβТ-клеток экспрессируется примерно в 5 раз
меньше молекул TCR, чем BCR на поверхности В-лимфоцитов
(30 000—40 000 TCR на клетку). Содержание комплексов CD3
примерно в 10 раз больше, чем TCR — около 300 000 молекул
на клетку, что свидетельствует о присутствии на мембране
молекул CD3, не связанных с TCR;
Помимо TCR—CD3 зрелые Т-клетки экспрессируют молекулы
CD2, CD5, CD7;
Наиболее важные в функциональном отношении мембранные
молекулы Т-клеток — корецепторы CD4 и CD8, служащие
маркерами основных субпопуляций αβТ-клеток, а также
костимулирующая молекула CD28, экспрессируемая на
большинстве (около 80%) Т-клеток;
Для наивных (не контактировавших с антигеном) αβТ-клеток
характерен высокий уровень экспрессии селектина L (CD62L)
и хемокинового рецептора CCR7. Эти молекулы определяют
пути миграции Т-клеток;
На Т-клетках содержатся также β1- и β2-интегрины (особенно
LFA-1 и VLA-4) и рецепторы для цитокинов (для IL-7, IL-1, IL-2,
IL-4, IL-15 и др.). Маркером наивных Т-клеток, отличающим их
от клеток памяти, служит полноразмерная форма молекулы
CD45 — CD45RA.
4.
Т-лимфоциты — активно рециркулирующие клетки;
Основное место локализации Т-лимфоцитов в
лимфоидных органах — тимусзависимые зоны
(паракортикальные зоны лимфатических узлов и
параартериальные муфты селезенки.) Вне Т-зон Тлимфоциты непосредственно соседствуют с В-клетками;
αβТ-лимфоциты диффузно распределены в
эпителиальных пластах барьерных тканей (где они
численно преобладают над γδТ-клетками) — слизистых
оболочках и эпидермисе. Их выявляют также в
соединительнотканных отделах барьерных тканей —
субмукозе и дерме;
Т-лимфоциты относят к долгоживущим клеткам. Срок
жизни различных субпопуляций наивных αβТлимфоцитов составляет месяцы и годы;
Для выживания Т-лимфоцитов необходим IL-7, а также
сигналы от TCR, при распознавании ими молекул МНС и
аутологичных пептидов.
5. αβТ-лимфоциты
6. СD8+ Т-лимфоциты (ЦТЛ)
Имеют цитотоксическую направленность
против изменённых клеток собственного
организма (инфицированные вирусом клетки,
опухолевые клетки-мишени);
Распознают антиген в ассоциации с молекулами
МНС I класса (феномен двойного
распознавания.)
7. CD4+ Т-лимфоциты (Th)
Поставляют вспомогательные сигналы при
активации В-лимфоцитов и макрофагов;
Взаимодействие Т-хелперов с дендритными
клетками служит пусковым событием Тзависимого иммунного ответа;
Вступают в связь с инвариантной частью
молекулы МНС II.
8. Регуляторные Т-клетки (Treg) (CD4+CD25+FoxP3+)
Некоторые CD4+ Т-клетки, экспрессирующие
внутриклеточный фактор FoxP3 и мембранные
молекулы CD25 и CTLA-4 (CD152), образуют
самостоятельную субпопуляцию естественных
регуляторных Т-клеток (Тreg). Их основная
функция — предотвращение реакции других Тклеток на аутоантигены, а также ограничение
(супрессия) любых форм иммунного ответа.
9. γδT-лимфоциты
10. γδT-лимфоциты
Вариабельность γδTCR ограничена и спектр
антигенов, распознаваемых γδТ-клетками, узок.
γδТ-клетки распознают антиген независимо от
молекул МНС. Поэтому корецепторы CD4 и CD8 не
обязательно присутствуют на их поверхности.
Сосредоточены преимущественно в барьерных
тканях.
Среди γδТ-лимфоцитов выделяют субпопуляцию
клеток, экспрессирующих гомодимерный вариант
молекулы CD8 — CD8αα (CD8αα+ γδТ-клетки.)
11. Этапы дифференцировки Т-лимфоцитов
12. Этапы дифференцировки Т-лимфоцитов
13. ELP: CD34+SCA-l+CD117(c-Kit)+Flt-3+ CCR9+ CD4lo
Sca-1 — антиген
стволовых клеток
(Stem cell antigen);
с-Kit — лиганд
фактора стволовых
клеток SCF (Stem cell
factor)
CD34 — используется
в качестве
идентификационного
для выявления
стволовых клеток и
их ближайших
потомков.
14. DN1
Тимоциты интенсивно пролиферируют (до 10
делений), что приводит к 1000-кратному
увеличению их числа;
Длительность периода — 10 сут;
Основной ростовой фактор на этом этапе — SCF (CKit лиганд);
Рецептор для IL-7 экспрессирован слабо или вообще
не экспрессирован;
Факторами дифференцировки на этом этапе
развития служат Wnt и Hedgehog, а также Notch,
коммутирующий клетки-предшественники к
дифференцировке в Т-лимфоциты.
15. DN2
Тимоциты перемещаются в сторону наружных
слоев коры по градиенту CXCL12, секретируемого
эпителиальными клетками наружной коры;
Длительность стадии — 2 сут;
Клетки продолжают пролиферировать уже
преимущественно под действием IL-7 и в меньшей
степени — SCF;
Сигналы от Notch коммиттируют Т-клетки к
дифференцировке в различные линии (γδ или αβ) и
экспрессируется ген, кодирующий pTCRα, — знак
выбора αβ-направления развития клетки.
16. DN3
Запускается перестройка V-генов TCR;
Для раннего развития Т-клеток необходима экспрессия
генов Notch, Runx-1, GATA-3, Ikaros, Gfil, c-Myb, PU1, E2A/HEB,
TCF-1;
Для запуска перестройки V-генов TCR наиболее важна
передача сигнала, осуществляемая Notch-1 на стадиях
DN2 и DN3. Рецептор для этого фактора экспрессируется
тимоцитами на указанных стадиях развития. Лиганды
Notch-рецептора — молекулы Delta и Jagged — находятся
на поверхности эпителиальных клеток тимуса.
Цитокины играют пределенную роль в индукции
перестройки V-генов, особенно IL-7, который особенно
важен для запуска перестройки V-гена γ-цепи. Еще более
существенна роль IL-7 в качестве фактора выживания,
индуцирующего экспрессию антиапоптотического
фактора Всl-2;
На заключительном этапе фазы DN3 на клетках
экспрессируется корецептор CD4 (стадия ISP.)
17. Роль Notch-рецепторов в развитии Т-клеток в тимусе
18. Реаранжировка V-генов TCR
19.
20. Выбор пути αβ/γδ-дифференцировки Т-клеток
21. DP
Тимоциты приобретают фенотип CD44-CD25- одновременно с
экспрессией корецепторов CD4 и CD8 (точнее, экспрессией
молекулы CD8 в дополнение к уже присутствующей на
мембране молекулы CD4).
Продолжительность стадии 1,5—2 сут;
Перестройка V-гена α-цепи;
Все это время не прекращаются экспрессии генов,
кодирующих RAG-1, RAG-2 и TdT;
После перестройки V-генов обеих цепей происходит сборка их
продуктов и экспрессия «зрелого» TCR на поверхности клетки;
Зрелый TCR экспрессируется в комплексе с CD3 и ζ-цепями;
Появление на поверхности тимоцита зрелого рецептора
служит сигналом, необходимым для прекращения экспрессии
Т-клеткой генов RAG-1 и RAG-2 и для поддержания ее
жизнеспособности;
Плотность экспрессии TCR на поверхности тимоцитов сначала
невелика;
Выживаемость клеток в этот период обеспечивается
взаимодействием молекулы семейства TNF — CD70 тимоцита
и молекулы семейства TNFR — CD27 эпителиальной клетки.
22. Селекция тимоцитов и формирование субпопуляций CD4+ и CD8 + клеток
Положительная селекция – обеспечивает отбор
только тех тимоцитов, которые экспрессируют
TCR, обладающие сродством к молекулам МНС.
Положительной селекции подвергаются
дважды положительные тимоциты фенотипа
CD4+CD8+CD3loCD27-;
Отрицательная селекция – ограничивает
степень агрессивности Т-клеток в отношении
собственных молекул. Тимоциты представлены
дважды положительными клетками или
«полузрелыми» моноположительными
клетками фенотипа CD4+/CD8+ CD24hi.
23. Положительная селекция
«по умолчанию»
24. Отрицательная селекция
25. Дифференцировка CD4 + и CD8 + тимоцитов
Дифференцировка Т-клеток регулируется как
внутренними (дифференцировочные факторы), так
и внешними (сигналы, генерируемые при контактах
между клетками) стимулами;
Факторы Notch не оказывают решающего влияния
на выбор пути дифференцировки;
Для дифференцировки Т-лимфоцитов в CD8+ клетки
нужен сигнал большей интенсивности, чем для
дифференцировки CD4+ Т-клетки;
Сочетанная экспрессия в клетках факторов Th-POK
и GATA-3 направляет Т-клетку по CD4+-пути;
Экспрессия факторов Тох и Runx3 направляет Тклетку по CD8+ пути.
26. Дифференцировка CD4 + и CD8 + тимоцитов
27. Проверка специфичности корецептора при дифференцировке CD4+ и CD8+ Т-лимфоцитов (селекционная модель).
28.
Созревшие тимоциты довольно долго (7—14 сут) не
покидают тимус, пребывая преимущественно в
наружных слоях мозгового слоя;
Завершается формирование аутотолерантности и Тклетки приобретают свойства, важные для поддержания
их жизнеспособности вне тимуса:
Вследствие активации сиалилтрансфераз происходит
сиалирование мембранных гликопротеинов,
защищающее лимфоциты от поглощения макрофагами;
Созревшие Т-лимфоциты начинают экспрессировать
набор мембранных молекул адгезии и рецепторов для
хемокинов, необходимые для направленной миграции
клеток в специализированные участки иммунной
системы и для последующей рециркуляции;
Под влиянием фактора KLF2 (Kruppel-like factor 2) на
тимоцитах экспрессируются селектин L (CD62L), β7интегрины, хемокиновый рецептор CCR7 и рецептор SIP1 для сфингозин-1-фосфата (играет основную роль в
эмиграции зрелых Т-клеток из тимуса в кровяное русло.)
29. Регуляторные Т-клетки (Treg)
Имеют мембранный фенотип CD4+CD25hiCTLA-4+GITR+PD-l+
Сильно экспрессируют α-цепь рецептора для IL-2 (CD25);
На их поверхности присутствуют супрессорные аналоги
костимулирующих молекул CTLA-4 и PD-1 и представитель
семейства TNFR — GITR (Glucocorticoid-induced TNFRrelated);
Супрессорная активность CD4+ регуляторных Т-клеток
связана с транскрипционным фактором FOXP3 (скурфин)
Сохраняют способность распознавать аутоантигены с
высокой степенью сродства (не проходят отрицательную
селекцию);
Устойчивы к апоптозу;
Приобретают способность вырабатывать суперссорные
цитокины (IL-10, трансформирующй фактор роста β).
30. Регуляторные Т-клетки (Treg)
31. Экспрессия генов CD25 и FOXP3 как ключевые события дифференцировки Treg
32. Спасибо за внимание.
Источник

ЛЕКЦИЯ № 4 Дифференцировка Т- и В-лимфоцитов
 малодифференцированный клеточный элемент, способный к самоподдержанию, пролиферации и дифференцировке")
Полипотентная гемопоэтическая стволовая клетка (ГСК) малодифференцированный клеточный элемент, способный к самоподдержанию, пролиферации и дифференцировке во все клетки крови.
,")
• ГСК локализуются в красном костном мозге (концентрация до 1% среди мононуклеарных клеток), • печени эмбриона (до 1%), • ГСК циркулируют в пуповинной крови (до 1%), • периферической крови (около 0, 1% от всех ядерных клеток)
. Родоначальная стволовая кроветворная")
Этапы развития ГСК • • Полипотентная стволовая кроветворная клетка (stem cell). Родоначальная стволовая кроветворная клетка (progenitor cell) – ограничена в самоподдержании, но интенсивно пролиферирует. Имеет ограниченное число делений и ограниченный потенциал, коммитированы к дифференцировке, по крайней мере, в двух направлениях (лимфоидном и миелоидном). Клетка предшественник (precursor cell) – коммитирована только в один тип клеток крови (лимфоциты, гранулоциты, моноциты и т. д. ). Зрелые клетки (Т-, В-лимфоциты, эритроциты, моноциты и т. п. ).

Лимфоидная родоначальная клетка ГСК самоподдержание В-лимфоцит Предшественник В-лимфоцитов Т-лимфоцит NK Предшественник Т-лимфоцитов Миелоидная родоначальная клетка

Treg самоподдержани е ТК? эоинофил базофил тромбоцит эритроцит

Клональный принцип организации лимфоцитов Клон лимфоцитов – группа лимфоцитов, имеющих антигенраспознающие рецепторы одной специфичности, т. е. способные распознавать одну АГ детерминанту. 1 КЛОН = 1 АГ-распознающий РЕЦЕПТОР! В иммунной системе формируется множество клонов лимфоцитов способных распознавать огромное количество вариантов антигенных молекул, составляющих антигенный репертуар.

Дифференцировка В-лимфоцитов

Субпопуляции В-клеток
 M")
Строение В-клеточного рецептора (BCR – “B-cell receptor”) M
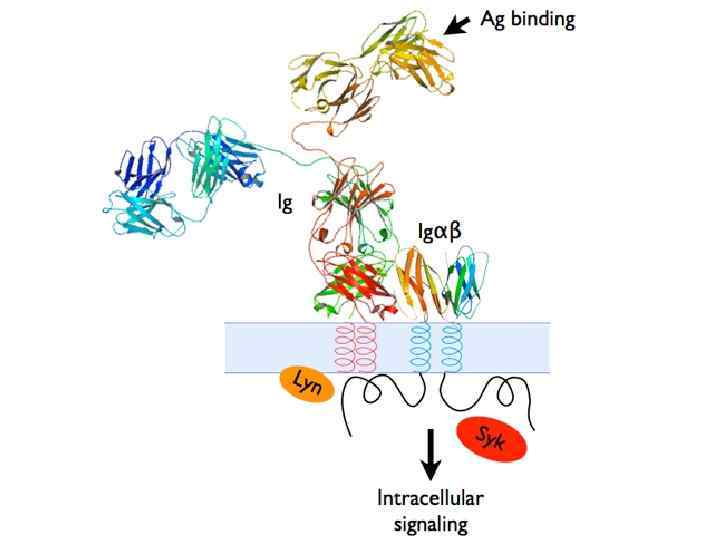
 сигнала от BCR")
Молекула BCR имеет слишком короткий цитоплазматический хвост, поэтому для трансдукции (передачи) сигнала от BCR к ядру клетки используются вспомогательные молекулы, такие как CD 79 и другие (CD 19, CD 20, CD 21).
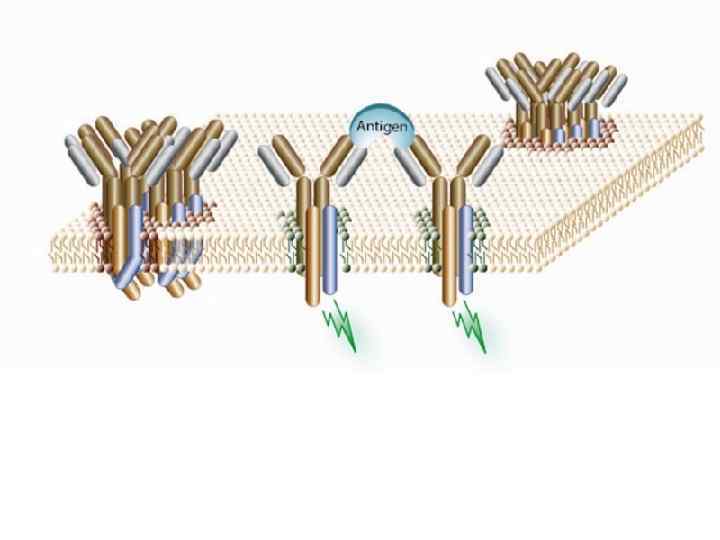

Основным процессом дифференцировки В-лимфоцитов является формирование антигенраспознающего В-клеточного рецептора!

Легкие цепи, входящие в состав BCR, могут быть двух видов: κиλ Их строение кодируется генетическими локусами, расположенными во 2 и 22 хромосомах Тяжелые цепи (5 видов, соответствующие классам иммуноглобулинов: μ, γ, δ, ε, α) кодируются в едином локусе 14 хромосомы. Это – зародышевые гены иммуноглобулинов.

Локализация генов иммуноглобулинов у человека и мыши

В общем виде каждая группа зародышевых генов состоит из 4 -х генных сегментов: ВАРИАБЕЛЬНЫЙ (V), РАЗНООБРАЗНЫЙ (D), СОЕДНИТЕЛЬНЫЙ (J) КОНСТАНТНЫЙ (С)

Организация зародышевых генов
 комплементарных различным видам АГ, во время дифференцировки происходит")
Для формирования многообразия вариабельных доменов (V-домены) комплементарных различным видам АГ, во время дифференцировки происходит перестройка (реаранжировка) зародышевых генов, в результате которой формируется окончательный вид генов, кодирующих белковую структуру BCR и иммуноглобулинов.
 зародышевых генов")
Реаранжировка (перестройка) зародышевых генов

С-гены тяжелой цепи BCR
.")
Для перегруппировки генов H- и L-цепей требуются ферменты рекомбиназы RAG-1 и RAG-2 (recombinationactivating genes). Перегруппировка представляет собой вырезание отдельных фрагментов ДНК и сшивку оставшихся частей по принципу образования петель ДНК.

Комбинаторное разнообразие антиген-связывающих участков

В результате реаранжировки генов BCR образуется более 2, 5 миллионов вариантов антиген-распознающих рецепторов.

Этапы дифференцировки B-лимфоцитов

Пре В-клеточный рецептор

Отрицательная селекция В-лимфоцитов в костном мозгу: реакции незрелых клеток на распознавание аутоантигена – индукция анергии, редактирование V-генов и апоптоз
")
Антиген-зависимый этап дифференцировки В-лимфоцитов (гуморальный иммунный ответ)

Дифференцировка В-лимфоцита в плазматическую клетку Плазматическая клетка B-лимфоцит Утрата экспрессии генов В-клетки Блокада клеточного цикла после плазмобласта Увеличение размера, расширение цитоплазмы, развитие шероховатого ЭПР

Дифференцировка Т-лимфоцитов
")
Строение T-клеточного рецептора (TCR – “T-cell receptor”)

3 D модель T-клеточного рецептора
 TCR тесно соединяется")
TCR – гетеродимеры, и или и цепей ( Т-клетки и Т-клетки) TCR тесно соединяется с CD 3 – комплексом полипептидных цепей, необходимых для трансдукции сигнала
 и CD 8 (αβ – гетеродимер")
Схематическая структура CD 4 (4 подобных Ig домена) и CD 8 (αβ – гетеродимер и αα- гомодимер)

В отличие от BCR, распознающего АГ в нативном виде, TCR связывает и распознает АГ только в составе молекул MHC. CD 4 и CD 8 классические корецепторы, распознающие комплекс «пептид-МНС» и участвующие в сигнальной трансдукции. Внеклеточные домены CD 4 и CD 8 связываются с консервативными МНС молекулами на АПК. CD 4 связывается с МНС класса II, а CD 8 – с МНС класса I

Распознавание антигена в составе молекул МНС I и II классов
 (CD 4+) Функциональная субпопуляция Т клеток, которые способствуют генерации цитотоксических Т лимфоцитов")
Т-хелперы (Th) (CD 4+) Функциональная субпопуляция Т клеток, которые способствуют генерации цитотоксических Т лимфоцитов и участвуют в межклеточной кооперации с В-клетками, активируя их в направлении синтеза антител. Th распознают антиген в ассоциации с молекулами MHC класса II (феномен двойного распознавания)
 Клетки-киллеры, способные убивать инфицированные вирусом клетки, опухолевые клеткимишени, несущие")
Цитотоксические Т клетки (СD 8+) Клетки-киллеры, способные убивать инфицированные вирусом клетки, опухолевые клеткимишени, несущие на поверхности антигенные пептиды в комплексе с молекулами MHC класса I (феномен двойного распознавания, но для эффекторной функции)
 (CD 4+CD 25+Fox. P 3+) Т-клетки, зависимые от тимуса, регулирующие (супрессирующие)")
Регуляторные Т-клетки (Treg) (CD 4+CD 25+Fox. P 3+) Т-клетки, зависимые от тимуса, регулирующие (супрессирующие) функцию других клеток иммунной системы

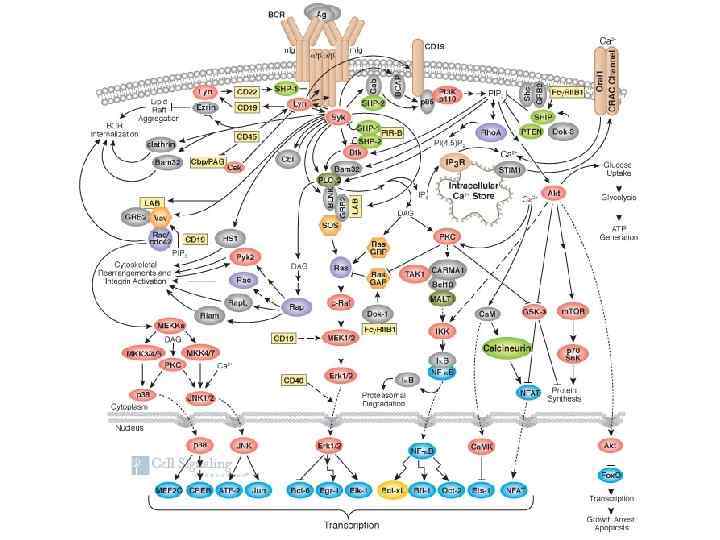
Основным процессом дифференцировки Т-лимфоцитов, также как и у В-лимфоцитов, является формирование многообразия антигенраспознающих Т-клеточных рецепторов

Локализация генов ТКР у человека

Реаранжировка генов по принципу образования петли ДНК

Циркулярные образования ДНК в Т-клетках, недавно покинувших тимус

Этапы дифференцировки Т-лимфоцитов

Положительная селекция тимоцитов

Отрицательная селекция тимоцитов

Положительная и отрицательная селекция тимоцитов

Спасибо за внимание.
Источник
Вы можете изучить и скачать доклад-презентацию на
тему Дифференцировка Т-лимфоцитов.
Презентация на заданную тему содержит 32 слайдов. Для просмотра воспользуйтесь
проигрывателем,
если материал оказался полезным для Вас – поделитесь им с друзьями с
помощью социальных кнопок и добавьте наш сайт презентаций в закладки!
Слайды и текст этой презентации
Слайд 1
Описание слайда:
Дифференцировка Т-лимфоцитов
Выполнила
Казьмина Н.А
3.4.12б
Слайд 2
Слайд 3 от общего числа лимфоцитов; абсолютное — (950—2100)х109 клеток в литре;
На поверхности αβТ-клеток экспрессируется примерно в 5 раз меньше молекул TCR, чем BCR на поверхности В-лимфоцитов (30 000—40 000 TCR на клетку). Содержание комплексов CD3 примерно в 10 раз больше, чем TCR — около 300 000 молекул на клетку, что свидетельствует о присутствии на мембране молекул CD3, не связанных с TCR;
Помимо TCR—CD3 зрелые Т-клетки экспрессируют молекулы CD2, CD5, CD7;
Наиболее важные в функциональном отношении мембранные молекулы Т-клеток — корецепторы CD4 и CD8, служащие маркерами основных субпопуляций αβТ-клеток, а также костимулирующая молекула CD28, экспрессируемая на большинстве (около 80%) Т-клеток;
Для наивных (не контактировавших с антигеном) αβТ-клеток характерен высокий уровень экспрессии селектина L (CD62L) и хемокинового рецептора CCR7. Эти молекулы определяют пути миграции Т-клеток;
На Т-клетках содержатся также β1- и β2-интегрины (особенно LFA-1 и VLA-4) и рецепторы для цитокинов (для IL-7, IL-1, IL-2, IL-4, IL-15 и др.). Маркером наивных Т-клеток, отличающим их от клеток памяти, служит полноразмерная форма молекулы CD45 — CD45RA.")
Описание слайда:
Относительное содержание Т-лимфоцитов в крови составляет в среднем около 73% (55-85%) от общего числа лимфоцитов; абсолютное — (950—2100)х109 клеток в литре;
На поверхности αβТ-клеток экспрессируется примерно в 5 раз меньше молекул TCR, чем BCR на поверхности В-лимфоцитов (30 000—40 000 TCR на клетку). Содержание комплексов CD3 примерно в 10 раз больше, чем TCR — около 300 000 молекул на клетку, что свидетельствует о присутствии на мембране молекул CD3, не связанных с TCR;
Помимо TCR—CD3 зрелые Т-клетки экспрессируют молекулы CD2, CD5, CD7;
Наиболее важные в функциональном отношении мембранные молекулы Т-клеток — корецепторы CD4 и CD8, служащие маркерами основных субпопуляций αβТ-клеток, а также костимулирующая молекула CD28, экспрессируемая на большинстве (около 80%) Т-клеток;
Для наивных (не контактировавших с антигеном) αβТ-клеток характерен высокий уровень экспрессии селектина L (CD62L) и хемокинового рецептора CCR7. Эти молекулы определяют пути миграции Т-клеток;
На Т-клетках содержатся также β1- и β2-интегрины (особенно LFA-1 и VLA-4) и рецепторы для цитокинов (для IL-7, IL-1, IL-2, IL-4, IL-15 и др.). Маркером наивных Т-клеток, отличающим их от клеток памяти, служит полноразмерная форма молекулы CD45 — CD45RA.
Слайд 4 Вне Т-зон Т-лимфоциты непосредственно соседствуют с В-клетками;
αβТ-лимфоциты диффузно распределены в эпителиальных пластах барьерных тканей (где они численно преобладают над γδТ-клетками) — слизистых оболочках и эпидермисе. Их выявляют также в соединительнотканных отделах барьерных тканей — субмукозе и дерме;
Т-лимфоциты относят к долгоживущим клеткам. Срок жизни различных субпопуляций наивных αβТ-лимфоцитов составляет месяцы и годы;
Для выживания Т-лимфоцитов необходим IL-7, а также сигналы от TCR, при распознавании ими молекул МНС и аутологичных пептидов.")
Описание слайда:
Т-лимфоциты — активно рециркулирующие клетки;
Основное место локализации Т-лимфоцитов в лимфоидных органах — тимусзависимые зоны (паракортикальные зоны лимфатических узлов и параартериальные муфты селезенки.) Вне Т-зон Т-лимфоциты непосредственно соседствуют с В-клетками;
αβТ-лимфоциты диффузно распределены в эпителиальных пластах барьерных тканей (где они численно преобладают над γδТ-клетками) — слизистых оболочках и эпидермисе. Их выявляют также в соединительнотканных отделах барьерных тканей — субмукозе и дерме;
Т-лимфоциты относят к долгоживущим клеткам. Срок жизни различных субпопуляций наивных αβТ-лимфоцитов составляет месяцы и годы;
Для выживания Т-лимфоцитов необходим IL-7, а также сигналы от TCR, при распознавании ими молекул МНС и аутологичных пептидов.
Слайд 5
Описание слайда:
αβТ-лимфоциты
Слайд 6
Имеют цитотоксическую направленность против изменённых клеток собственного организма (инфицированные вирусом клетки, опухолевые клетки-мишени);
Распознают антиген в ассоциации с молекулами МНС I класса (феномен двойного распознавания.)")
Описание слайда:
СD8+ Т-лимфоциты (ЦТЛ)
Имеют цитотоксическую направленность против изменённых клеток собственного организма (инфицированные вирусом клетки, опухолевые клетки-мишени);
Распознают антиген в ассоциации с молекулами МНС I класса (феномен двойного распознавания.)
Слайд 7
Поставляют вспомогательные сигналы при активации В-лимфоцитов и макрофагов;
Взаимодействие Т-хелперов с дендритными клетками служит пусковым событием Т-зависимого иммунного ответа;
Вступают в связь с инвариантной частью молекулы МНС II.")
Описание слайда:
CD4+ Т-лимфоциты (Th)
Поставляют вспомогательные сигналы при активации В-лимфоцитов и макрофагов;
Взаимодействие Т-хелперов с дендритными клетками служит пусковым событием Т-зависимого иммунного ответа;
Вступают в связь с инвариантной частью молекулы МНС II.
Слайд 8 (CD4+CD25+FoxP3+)
Некоторые CD4+ Т-клетки, экспрессирующие внутриклеточный фактор FoxP3 и мембранные молекулы CD25 и CTLA-4 (CD152), образуют самостоятельную субпопуляцию естественных регуляторных Т-клеток (Тreg). Их основная функция — предотвращение реакции других Т-клеток на аутоантигены, а также ограничение (супрессия) любых форм иммунного ответа.")
Описание слайда:
Регуляторные Т-клетки (Treg) (CD4+CD25+FoxP3+)
Некоторые CD4+ Т-клетки, экспрессирующие внутриклеточный фактор FoxP3 и мембранные молекулы CD25 и CTLA-4 (CD152), образуют самостоятельную субпопуляцию естественных регуляторных Т-клеток (Тreg). Их основная функция — предотвращение реакции других Т-клеток на аутоантигены, а также ограничение (супрессия) любых форм иммунного ответа.
Слайд 9
Описание слайда:
γδT-лимфоциты
Слайд 10")
Описание слайда:
γδT-лимфоциты
Вариабельность γδTCR ограничена и спектр антигенов, распознаваемых γδТ-клетками, узок.
γδТ-клетки распознают антиген независимо от молекул МНС. Поэтому корецепторы CD4 и CD8 не обязательно присутствуют на их поверхности.
Сосредоточены преимущественно в барьерных тканях.
Среди γδТ-лимфоцитов выделяют субпопуляцию клеток, экспрессирующих гомодимерный вариант молекулы CD8 — CD8αα (CD8αα+ γδТ-клетки.)
Слайд 11
Описание слайда:
Этапы дифференцировки Т-лимфоцитов
Слайд 12
Описание слайда:
Этапы дифференцировки Т-лимфоцитов
Слайд 13+Flt-3+ CCR9+ CD4lo
Sca-1 — антиген стволовых клеток (Stem cell antigen);
с-Kit — лиганд фактора стволовых клеток SCF (Stem cell factor)
CD34 — используется в качестве идентификационного для выявления стволовых клеток и их ближайших потомков.")
Оп