Циркуляция лимфоцитов регулируется чем

Лимфоциты постоянно рециркулируют из крови в ткани, и обратно. Рециркуляцией называют процесс перехода лимфоцитов из кровяного русла в органы, оттуда в лимфу и вновь в кровоток. При этом клетки, покинувшие конкретный лимфатический узел, возвращаются в любой узел или селезенку, что и обеспечивает перемешивание рециркулирующих лимфоцитов.
Лимфоциты переходят из кровотока в лимфоидные образования обычные посткапиллярные венулы, однако, этот переход обычно осуществляется преимущественно через специализированные участки венозного русла – венулы с высоким эндотелием. В лимфоузлах эти сосуды находятся главным образом в паракортикальной области и иногда в корковой. Вместе с тем часть лимфоцитов, а в первую очередь Т-клетки, поступают в региональный лимфатический узел из дренируемой им области по приносящим лимфатическим сосудам.
Из лимфатических узлов лимфоциты возвращаются в кровоток по приносящим лимфатическим сосудам через грудной поток в краниальную полую вену. Ежечасно в рециркуляцию вовлекается 1-2% лимфоцитов, большинство зрелых лимфоцитов поступает из органов в циркуляцию и обратно, причем, время, соответствующее половине срока их однократного пребывания в циркуляции, составляет до 30 минут. В итоге этот процесс позволяет множеству антигенспецифичных лимфоцитов встретиться с соответствующими антигенами, проникшими в периферические лимфоидные органы. Особая важность рециркуляции становится очевидной, если учесть, что лимфоциты моноспецифичны и лишь ограниченное число лимфоцитов способно распознавать каждый конкретный антиген.
В норме рециркуляция лимфоцитов через лимфатические узлы происходит постоянно. Однако, если в лимфатический узел, ранее сенсибилизированный к конкретному антигену, повторно попадает антиген, то рециркуляция прекращается примерно на 24 часа. Временная остановка рециркуляции обусловлена избирательной задержкой антигенспецифичных лимфоцитов в лимфоузлах, дренирующих место проникновения антигена. Лимфоциты уже не рециркулируют, а остаются в участке встречи с антигеном.
Специфическая рециркуляция лимфоцитов происходит в лимфоидной ткани, ассоциированной со слизистыми оболочками. Лимфоциты этой ткани возвращаются в процессе рециркуляции образом в эту же ткань. Так, лимфоциты, стимулированные в пейеровых бляшках, проходят через региональные лимфатические узлы в кровоток, а затем возвращаются «домой» в собственную пластинку слизистой оболочки. Такая специфическая рециркуляция объясняется тем, что эти лимфоциты экспрессируют молекулы «возврата домой», которые связываются со специальными молекулами адгезии-адрессиинами на поверхности эндотелиальных клеток венул лимфоидной ткани слизистых оболочек, что и обеспечивает избирательную рециркуляцию. По этой же причине стимуляция антигеном в области слизистой оболочки в том или ином участке организма вызывает образование антител преимущественно в слизистых оболочках.
В определении направления миграции лимфоцитов и в преодолении барьеров между кровью и тканями основную роль играют хемотаксические сигналы и специфические межклеточные взаимодействия, основанные на взаимном распознавании мембранных структур клеток.
Источник
ЦИРКУЛЯЦИЯ ЛИМФЫ
Рис. 1. Афферентные лимфатические сосуды (АЛС) доставляют лимфу с лимфоцитами из соединительных тканей или периферических лимфатических узлов в субкапсулярный синус (СС). Оттуда лимфа проходит через промежуточные синусы (ПС) в мозговые синусы (МС), чтобы попасть в эфферентные лимфатические сосуды (ЭЛС). В районе ворот (В) лимфа из субкапсулярного синуса идет прямо в мозговые синусы. Лимфа также инфильтрирует лимфоидную ткань и медленно течет от коркового (KB) к мозговому веществу (MB). Лимфоциты покидают паренхиму лимфатического узла через мозговые синусы, достигая эфферентного лимфатического сосуда. Направление циркуляции лимфы (стрелки) регулируется клапанами (К).
КРОВОСНАБЖЕНИЕ ЛИМФАТИЧЕСКОГО УЗЛА
Рис. 2. Кровоснабжение каждого лимфатического узла происходит за счет маленькой мышечной артерии (А), которая входит в лимфатический узел через его ворота (В), разветвляясь затем на артериолы (Арт), идущие через мозговые тяжи (MT). Достигнув важнейшего коркового слоя (КС), артериолы распадаются на капиллярную сеть вокруг и внутри герминативных центров лимфоидных узелков (ЛиУ). Кровь затем собирается посткапиллярными венулами (ПсКВ), которые проходят радиально через внутренний слой коркового вещества (ВКВ), входят в мозговые тяжи и объединяются в большую вену (Be), которая покидает лимфатичекий узел через его ворота. Как отмечалось ранее, посткапиллярные венулы — это места, где лимфоциты покидают поток крови, чтобы войти в паренхиму лимфатического узла.
ЛИМФОЦИТАРНАЯ ЦИРКУЛЯЦИЯ (ЦИРКУЛЯЦИЯ ЛИМФОЦИТОВ)
Рис. 3. Лимфоцитарная циркуляция — это медленное движение лимфопоэтических клеток и лимфоцитов между различными лимфоидными органами и тканями посредством крови и лимфы.
Т- и В-лимфообразующие клетки появляются из лимфоидной клетки-предшественницы (ЛКП) костного мозга. Предшественники Т-лимфоцитов (Т) мигрируют через кровоток в тимус, где дифференцируются в зрелые иммунокомпетентные Т-лимфоциты в контакте с эпителиоретикулярными клетками тимуса, находясь в особом микроокружении этого органа.
Большинство В-лимфоцитов (В), формирующихся в костном мозге, поступает в кровоток, часть клеток-предшественниц В-лимфоцитов остается в костном мозге для поддержания популяции. Отсюда они мигрируют в аналог сумки Фабрициуса* (аппендикс, пейеровы бляшки и миндалины), где и происходит их созревание. Иммунокомпетентные В-лимфоциты развиваются из этих клеток митотическим путем. Из тимуса и аналога сумки Фабрициуса Т- и В-лимфоциты опять поступают в кровоток, затем покидают его, мигрируя в ткани. Лимфоциты собираются в лимфатических узлах после прохождения через афферентные лимфатические сосуды (АЛС).
Через эфферентные лимфатические сосуды (ЭЛС) лимфоциты опять проходят в кровоток и ткани, и цикл повторяется. Лимфоцитарная циркуляция требует несколько недель.
РЕЦИРКУЛЯЦИЯ ЛИМФОЦИТОВ
Рис. 4. Лимфоцитарная рециркуляция — это явление, параллельное циркуляции лимфоцитов.
Она состоит из быстрой миграции В- и Т-лимфоцитов из крови в ткани и лимфоидные органы и затем назад в кровь.
Стрелки указывают направление лимфо- и кровотока.
В-лимфоциты (светлые кружочки) и Т-лимфоциты (темные кружочки) входят в ткани (Тk) из кровеносных капилляров (Кап), чтобы быть собранными афферентными лимфатическими сосудами (АЛС) в лимфатических узлах (ЛУ). Здесь некоторые Т-лимфоциты заселяют тимус зависимые зоны, но большинство лимфоцитов продвигается через синусы в паренхиму, поступая затем через эфферентные лимфатические сосуды (ЭЛС) в грудной проток (ГП), посредством которого они попадают в кровоток (КГ). Отсюда лимфоциты распределяются по лимфоидным органам (тимус — Т, аналог сумки—АС; селезенка —С, лимфатические узлы — ЛУ) и тканям. Т-лимфоциты образуются в тимусе, они не рециркулируют через его паренхиму, а покидают орган через его лимфатические и кровеносные сосуды. В лимфатических узлах лимфоциты проходят через стенки посткапиллярных венул (ПсКВ) и поступают в лимфоидную ткань, мигрируя через нее, чтобы вновь накопиться в эфферентных лимфатических сосудах. Лимфоцитарная рециркуляция через кровь занимает около 0,6 ч, прохождение через селезенку—около 6 ч, а через лимфатические узлы 15—20 ч. Во время этого периода рециркуляции лимфоциты не делятся. Лимфобласты не рециркулируют.
Цель лимфоцитарной рециркуляции — дать возможность иммунокомпетентным лимфоцитам осуществлять постоянную защиту организма и информировать лимфоидные органы о наличии или отсутствии антигенов в теле. В присутствии антигенов часть лимфоцитов оседает в лимфоидных органах и начинает делиться, приводя в действие иммунную реакцию. Те, которые потеряны при циркуляции, компенсируются за счет медленной лимфоцитарной циркуляции посредством воспроизводства в костном мозге, тимусе, селезенке и других лимфоидных органах.
*Сумка Фабрициуса — лимфоэпителиальный орган, обнаруженный у птиц в форме клоакального дивертикула, который участвует в производстве иммунокомпетентных В-лимфоцитов. Сумка Фабрициуса не существует у млекопитающих и человека, но предполагают, что лимфоидные узелки в миндалинах, аппендиксе, пейеровых бляшках и других органах могут частично выполнять эту роль. Поэтому эти органы называют аналогом сумки Фабрициуса, или эквивалентом сумки.
Источник
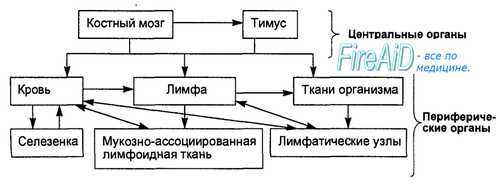
Лимфоидные органы — динамичные образования: лимфоциты непрерывно перемещаются в стромальном каркасе, покидают лимфоидные органы и вновь возвращаются в них. Этот круговорот лимфоцитов обозначают термином «рециркуляция». В процессе рециркуляции лимфоциты покидают лимфоидные органы с эфферентной лимфой (из селезенки они выходят с кровью). Через грудной лимфатический проток лимфоциты проникают в кровяное русло. Затем через посткапиллярные венулы клетки вновь проникают в лимфоидный орган (тот же или другой), внутри которого перемещаются в специфические места их локализации. Затем клетки вновь покидают орган и выходят в циркуляторное русло и т.д. В нелимфоидных органах лимфоциты практически не циркулируют.
Из сказанного следует, что выявляемые в крови лимфоциты находятся в ней недолго (примерно 30 мин), но в течение суток многократно (4—5 раз) покидают ее и вновь возвращаются. Таким образом, несмотря на то, что в крови лимфоциты составляют всего около 0,5% от общего числа клеток, их состояние достаточно точно отражает состояние популяции лимфоцитов в целом организме. Это очень существенно, поскольку именно лимфоциты крови человека служат наиболее обычным и доступным объектом изучения как в экспериментальных, так и в клинико-иммунологических исследованиях. Субпопуляционный состав лимфоцитов крови человека представлен в табл. 3.23.
Таблица 3.23. Нормальные показатели содержания в крови взрослых людей основных популяций и субпопуляций лимфоцитов (по С.В. Хайдукову, 2008)
| Клетки | Относительное содержание*, % | Число клеток в 1 л (М+m) |
| Лимфоциты (CD45+)** | 32+4 | 1,363—2,808х109 |
| В-клетки ^D3-CD19+) | 12+5 | 0,111—0,376х109 |
| Т-клетки (CD3+CD19-) | 73+12 | 0,946-2,079х109 |
| арТ-клетки (CD3+apTCR+) | 70,5+9,7 | 0,022—0,115х109 |
| yST-клетки (CD3+ySTCR+) | 4,6+2,8 | 0,924—1,964х109 |
| Т-хелперы (CD3+CD4+) | 45+10 | 0,576—1,336х109 |
| Т-киллеры (CD3+CD8+) | 27+8 | 0,372—0,974х109 |
| Регулятоные T-клетки (CD3+CD4+CD25hi) | 3,7+2,05 | 0,009—0,078х 109 |
| NKT-клетки (CD3+CD56+CD16+) | 3+3 | 0,007—0,165х109 |
| Т-хелперы наивные (CD3+CD4+CD45RA+) | 30+10 | 0,272—1,123х109 |
| Т-хелперы активированные/памяти (CD3+CD4+CD45R0+CD29+) | 15+10 | 0,068—0,702х109 |
| NK-клетки (CD45+CD56+СD16+) | 13+5 | 0,123—0,369х109 |
* Указано среднее арифметическое со стандартной ошибкой.
** Относительные и абсолютные количества лимфоцитов определяли от общего количества лейкоцитов.
Относительные и абсолютные количества субпопуляций Т-, В- и NK-клеток определяли от общего количества лимфоцитов.
Способность клеток находить «свое место» в организме называют хомингом. Существует две группы механизмов, задействованных в данном процессе, — контактные механизмы, обеспечиваемые молекулами адгезии, находящимися на поверхности лимфоцитов, и хемотаксис, определяющий направленность движения клеток, зависящую от наличия на их поверхности хемокиновых рецепторов.
Попав в сосудистую сеть лимфоидного органа, лимфоциты реагируют на хемокины, вырабатываемые клетками эндотелия в специализированном для миграции лимфоцитов отделе сосудов — посткапиллярных венулах.
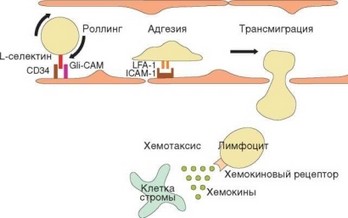
Рис. 3.74. Схема транссосудистой миграции лимфоцитов. Отражены основные стадии эмиграции лимфоцитов через высокий эндотелий посткапиллярных венул лимфоидных органов. Указаны молекулы адгезии, важные для осуществления этого процесса; отражена роль хемокинов в привлечении клеток
Только в посткапиллярных венулах лимфоидных (но не других) органов есть высокий эндотелий, образованный спонтанно активированными клетками (аналогичный высокий эндотелий может быть индуцирован в других органах только под влиянием провоспалительных факторов). Клетки этого эндотелия секретируют р-хемокин ССЫ9, распознаваемый рецептором CCR7. Этот рецептор экспрессируют на поверхности наивные лимфоциты как Т-, так и В-класса. Таким образом, указанные эндотелиальные клетки способны привлекать как Т-, так и В-лимфоциты.
Далее происходит трансмиграция лимфоцитов по механизму, аналогичному миграции лейкоцитов в очаг воспаления (отличия касаются конкретных молекул адгезии, определяющих направление миграции — рис. 3.74). Ключевую роль при этом играет начальный момент миграции, обусловленный взаимодействием мембранного L-cелектина (СD62L) лимфоцитов с адрессинами (GlyCAM, CD34, PSGL-1) эндотелиальных клеток. Именно наличие CD62L на клеточной мембране определяет способность наивных лимфоцитов мигрировать в лимфатические узлы и пейеровы бляшки, в связи с чем CD62L называют рецептором хоминга. Это слабое взаимодействие обусловливает феномен качения (перекатывания лимфоцита вдоль сосудистой стенки). Вслед за этим устанавливается более прочное взаимодействие между р2-интегрином лимфоцитов LFA-1 и его рецептором на эндотелиальной клетке — ICAM-1. Взаимодействие усиливается благодаря «активации» р2-интегрина (т.е. изменения конформации молекулы, в результате которого повышается его сродство к рецептору — см. раздел 2.3.1.2). Это обусловливает прочное прилипание лимфоцита к сосудистой стенке. Затем под влиянием хемотаксических сигналов, поступающих из ткани
лимфоидного органа, лимфоцит начинает мигрировать из сосуда между эндотелиальными клетками.
Вышедшие из сосуда клетки попадают в примыкающие к посткапиллярным венулам тимусзависимые зоны, стромальные и дендритные клетки которых секретируют хемокины CCL19 (ELC) и С^21 (SLC), распознаваемые рецептором CCR7 на лимфоцитах. До этого момента Т- и В-клетки перемещаются в одном и том же направлении. После того как лимфоцит попадает во внутреннюю среду органа, их пути расходятся. Т-клетки остаются в тимусзависимой зоне (в паракортексе лимфатических узлов, параартериальной муфте белой пульпы селезенки и Т-зонах пейеровых бляшек). В-клетки реагируют на хемокин CXCR13 (BLC), к которому они экспрессирут рецептор СХСR5. Этот хемокин секретируют стромальные клетки фолликулов, что и определяет направление миграции В-лимфоцитов. В-лимфоциты, не несущие СХСR5, не поступают в фолликулы и локализуются в прилежащей к фолликулу зоне. Механизмы, определяющие дальнейшее перемещение лимфоцитов из органа в эфферентную лимфу, пока не установлены.
В процессе рециркуляции отсутствует «привязка» хоминга к конкретному лимфатическому узлу или пейеровой бляшке. Так, лимфоцит, покинувший шейный лимфоузел, может затем проникнуть, например, в паховый лимфатический узел или в пейерову бляшку. Принципиально лишь то, что клетки не выходят из системы вторичных лимфоидных органов. Степень специфичности хоминга минимальна в селезенке, так как лимфоциты проникают в белую пульпу при излиянии крови в краевой зоне. Только последующее перемещение в Т- и В-зоны осуществляется по описанному выше механизму.
Все сказанное выше относится к наивным лимфоцитам. После активации лимфоцитов антигеном экспрессия молекул адгезии и хемокиновых рецепторов на их поверхности существенно изменяется. Это определяет иные пути миграции клеток после контакта с антигеном. Активированные лимфоциты и клетки памяти очень слабо экспрессируют L-селектин, что обусловливает низкую вероятность их попадания в лимфатические узлы. В то же время они приобретают способность мигрировать в очаги воспаления, расположенные в нелимфоидных органах, а также в барьерные ткани (рис. 3.75). Пути миграции активированных лимфоцитов и клеток

Рис. 3.75. Мембранный фенотип Т-клеток определяет пути их миграции. Указаны молекулы адгезии, определяющие различные направления миграции Т-клеток
памяти будут специально рассмотрены далее (см. раздел 3.6.5.4). Благодаря различной экспрессии рецепторов хоминга в разных тканях формируется несколько относительно автономных путей рециркуляции. Пути рециркуляции наивных лимфоцитов и клеток памяти могут пересекаться. Так, и наивные клетки, и клетки памяти, активированные в кишечнике, попадают в брыжеечные лимфатические узлы.
Интенсивность рециркуляции Т-лимфоцтов существенно выше, чем В-лимфоцитов. У человека через каждый лимфатический узел за сутки проходит 0,3х10п, через селезенку — 2,5х10п, а через кровь — 5х10п лимфоцитов. Длительность пребывания Т-клеток в кровотоке за 1 цикл рециркуляции — около 30 мин. Благодаря рециркуляции происходит постоянное «перемешивание» лимфоидных клеток с сохранением описанного выше порядка: наивные клетки, как правило, не покидают лимфоидной ткани, тогда как клетки памяти, наоборот, часто выходят за ее пределы, мигрируя, например, в эпителиальные органы и ткани, а также в очаги воспаления. Внутри лимфоидных органов клетки также занимают соответствующие места. Упорядоченность локализации лимфоцитов различных типов очень важна для развития иммунного ответа.
Источник
Оглавление темы “Иммунная система. Функции клеток иммунной системы. Т-лимфоциты. В-лимфоциты. Структура и функции органов иммунной системы.”: Иммунная система. Иммунная система человека. Неспецифический иммунитет. Специфический иммунный ответ. Рециркуляция лимфоцитов.Во внутренней среде организма присутствуют клетки и молекулы, которые обеспечивают защитную функцию (иммунитет). Часть из них относятся к факторам врожденного иммунитета, т. е. присутствуют в организме еще до встречи с каким-либо болезнетворным микроорганизмом или чужеродной молекулой. К таким факторам защиты относятся фагоцитирующие клетки крови и тканей, а также большие гранулярные лимфоциты, получившие название «натуральные (естественные) киллеры». Врожденными факторами защиты организма являются также молекулы, продуцируемые и секретируемые вышеназванными клетками, а также лимфоцитами, стромальными и другими клетками. К таким молекулам относятся: белки системы комплемента, медиаторы межклеточного взаимодействия — цитокины и др. Белки системы комплемента продуцируются постоянно и циркулируют в крови, большинство цитокинов начинают продуцироваться и секре-тироваться клетками только в ответ на сигнал активации (например, на контакт с микроорганизмом). Более эффективным способом защиты внутренней среды организма от проникающих в нее чужеродных агентов (антигенов) является специфический иммунный ответ, в результате которого организм приобретает дополнительные защитные механизмы: активированные клетки и продуцируемые ими молекулы. Защитное действие этих механизмов строго избирательно (специфично) в отношении того конкретного антигена (например, патогенного микроорганизма), контакт с которым вызвал иммунный ответ. Специфический иммунный ответ является функцией клеток и органов иммунной системы.
Главными клетками иммунной системы являются Т- и В-лимфоциты, которые рециркулируют в системе кровотока и лимфотока, постоянно перемещаясь из одних органов иммунной системы в другие, обладают способностью выходить в ткани для выполнения защитных функций (рис. 8.1). В защитных реакциях специфического иммунитета кроме Т- и В-лим-фоцитов участвуют фагоцитирующие клетки (гранулоциты, моноциты, макрофаги), «естественные киллеры, тучные клетки, эндотелиальные и эпителиальные клетки, которые играют роль вспомогательных, взаимодействуя с Т- и В-лимфоцитами. – Также рекомендуем “Происхождение ( образование ) клеток иммунной системы. Функции клеток иммунной системы. Лимфопоэз. Бурса Фабрициуса..” |
Источник